оНКЕГМНЕ:
йЮЙ ЯДЕКЮРЭ ПЮГЦНБНП ОНКЕГМШЛ Х ОПХЪРМШЛ
йЮЙ ЯДЕКЮРЭ НАЗЕЛМСЧ ГБЕГДС ЯБНХЛХ ПСЙЮЛХ
йЮЙ ЯДЕКЮРЭ РН, ВРН ДЕКЮРЭ МЕ УНВЕРЯЪ?
йЮЙ ЯДЕКЮРЭ ОНЦПЕЛСЬЙС
йЮЙ ЯДЕКЮРЭ РЮЙ ВРНАШ ФЕМЫХМШ ЯЮЛХ ГМЮЙНЛХКХЯЭ Я БЮЛХ
йЮЙ ЯДЕКЮРЭ ХДЕЧ ЙНЛЛЕПВЕЯЙНИ
йЮЙ ЯДЕКЮРЭ УНПНЬСЧ ПЮЯРЪФЙС МНЦ?
йЮЙ ЯДЕКЮРЭ МЮЬ ПЮГСЛ ГДНПНБШЛ?
йЮЙ ЯДЕКЮРЭ, ВРНАШ КЧДХ НАЛЮМШБЮКХ ЛЕМЭЬЕ
бНОПНЯ 4. йЮЙ ЯДЕКЮРЭ РЮЙ, ВРНАШ БЮЯ СБЮФЮКХ Х ЖЕМХКХ?
йЮЙ ЯДЕКЮРЭ КСВЬЕ ЯЕАЕ Х ДПСЦХЛ КЧДЪЛ
йЮЙ ЯДЕКЮРЭ ЯБХДЮМХЕ ХМРЕПЕЯМШЛ?

йЮРЕЦНПХХ:
юПУХРЕЙРСПЮюЯРПНМНЛХЪаХНКНЦХЪцЕНЦПЮТХЪцЕНКНЦХЪхМТНПЛЮРХЙЮхЯЙСЯЯРБНхЯРНПХЪйСКХМЮПХЪйСКЭРСПЮлЮПЙЕРХМЦлЮРЕЛЮРХЙЮлЕДХЖХМЮлЕМЕДФЛЕМРнУПЮМЮ РПСДЮоПЮБНоПНХГБНДЯРБНоЯХУНКНЦХЪпЕКХЦХЪяНЖХНКНЦХЪяОНПРрЕУМХЙЮтХГХЙЮтХКНЯНТХЪуХЛХЪщЙНКНЦХЪщЙНМНЛХЙЮщКЕЙРПНМХЙЮ
хЯґЯКЕґДНґБЮґМХЕ ЩКЕЙґРПХґВЕґЯЙНИ ЮЙґРХБґМНґЯРХ ЦНґКНБґМНґЦН ЛНГґЦЮ Х ПНКЭ ОЯХґУНґКНґЦХґВЕґЯЙНґЦН ЙНЛґОНґМЕМґРЮ Б ХМґДСЙґЖХХ хяя
|
|
оПНґАКЕґЛШ ХЯґЯКЕґДНґБЮґМХЪ ЛНГґЦЮ ВЕґКНґБЕґЙЮ, ЯНґЯРНЪґМХИ ЯНГґМЮґМХЪ, ЯНґНРґМНґЬЕґМХЪ ЛНГґЦЮ Х ОЯХґУХґЙХ √ ЪБґКЪґЧРґЯЪ НДґМХґЛХ ХГ ЯЮґЛШУ ГЮґУБЮґРШґБЮЧґЫХУ ГЮґДЮВ, ЙНґРНґПШЕ ЙНґЦДЮ-КХґАН ЯРЮґБХґКХЯЭ Б МЮСґЙЕ. хУ ХГСґВЕґМХЕ ГЮґЯРЮґБХґКН ЯСґЫЕґЯРґБЕМґМН ОЕґПЕґЯЛНРґПЕРЭ БГЦКЪґДШ МЮ ПЕґЮКЭґМНЯРЭ Х ОПХґПНґДС ВЕґКНґБЕґЙЮ.
йЮЙ ХГґБЕЯРґМН, щщц ЪБґКЪґЕРґЯЪ МЕґЯРЮґЖХНґМЮПґМШЛ ОПНґЖЕЯґЯНЛ, Х ДЮґФЕ ОПХ НРґЯСРґЯРґБХХ ЙЮґЙХУ-КХґАН ЪБґМШУ БНГґЛСґЫЮЧґЫХУ БМЕЬґМХУ ТЮЙґРНґПНБ Б МЕИ МЮґАКЧґДЮґЧРґЯЪ ЯСґЫЕґЯРґБЕМґМШЕ ХГґЛЕґМЕґМХЪ Б БХґДЕ ЯХМґУПНґМХґГЮґЖХХ, ДЕґЯХМґУПНґМХґГЮґЖХХ, БПЕґЛЕМґМШУ ЮЯХЛґЛЕРґПХИ, НАСґЯКНБґКЕМґМШУ ЯОНМґРЮМґМШґЛХ ЙНґКЕґАЮґМХЪґЛХ СПНБґМЪ ТСМЙґЖХНґМЮКЭґМНИ ЮЙґРХБґМНґЯРХ, НЯНґАЕМґМНґЯРЪґЛХ ОЯХґУХґВЕґЯЙНИ ЮЙґРХБґМНґЯРХ БН БПЕґЛЪ ПЕґЦХґЯРґПЮґЖХХ [151, 173].
б НАШВґМНЛ ЯНґЯРНЪґМХХ ЯНГґМЮґМХЪ ЯСґЫЕґЯРґБСґЕР ЯАЮґКЮМґЯХґПНґБЮМґМНЯРЭ БМСРґПХґЖЕМґРПЮКЭґМШУ БГЮХґЛНґНРґМНґЬЕґМХИ, Б РН БПЕґЛЪ ЙЮЙ Б хяя ОПНґХЯґУНґДХР ПЮЯґОЮД ТСМЙґЖХНґМЮКЭґМШУ ЯБЪґГЕИ, ВРН ОПНґЪБґКЪґЕРґЯЪ АНКЭґЬНИ МЕґСЯґРНИґВХґБНґЯРЭЧ МЕИґПНґДХґМЮґЛХґВЕґЯЙХУ ОПНґЖЕЯґЯНБ Х ПЮГґПСґЬЕґМХґЕЛ ╚ЛНґГЮХґЙХ╩ ЙНПґЙНґБН-ОНДґЙНПґЙНґБШУ НРґМНґЬЕґМХИ. оПХ ХГґЛЕґМЕґМХХ тя ЛНГґЦЮ ЯАЮґКЮМґЯХґПНґБЮМґМНЯРЭ ЙНПґЙНґБН-ОНДґЙНПґЙНґБШУ БГЮХґЛНґНРґМНґЬЕґМХИ МЮґПСґЬЮґЕРґЯЪ, ВРН ЯНґОПНґБНґФґДЮґЕРґЯЪ ХГґЛЕґМЕґМХґЕЛ ОЮРґРЕПґМНБ щщц.
тНґМНґБНґЛС ОЕґПХНґДС ДШґУЮґРЕКЭґМНИ ЯЕЯґЯХХ ЯНґНРґБЕРґЯРґБНґБЮґКЮ ОНґКХґЛНПТґМЮЪ ЮЙґРХБґМНЯРЭ НРґМНґЯХґРЕКЭґМН МХГґЙНИ ЮЛґОКХґРСґДШ Я МЮґКХґВХґЕЛ ЮКЭґТЮ-, АЕґРЮ-, РЕґРЮ- Х ДЕКЭґРЮ- БНКМ. пЕґКЮЙґЯЮґЖХЪ ОПХґБНґДХґКЮ Й СЯХґКЕґМХЧ ЛЕДґКЕМґМН-БНКґМНґБНИ ЮЙґРХБґМНґЯРХ Б МЕґЯЙНКЭґЙН ПЮГ Х ЯНґОПНґБНґФґДЮґКЮЯЭ ТНПґЛХґПНґБЮґМХґЕЛ ТНґЙСґЯНБ РЕґРЮ- Х ДЕКЭґРЮ-ПХРґЛЮ. мЮХґАНґКЕЕ РХґОХВґМНИ ХУ КНґЙЮґКХґГЮґЖХґЕИ ЪБґКЪґКХЯЭ, ЙЮЙ ОПЮґБХґКН, ОЕґПЕДґМХЕ КНАґМШЕ ДНґКХ, НДґМЮґЙН Б ЖЕґКНЛ ПЪґДЕ ЯКСґВЮґЕБ НРґЯСРґЯРґБНґБЮК ГЮґЙНґМНґЛЕПґМШИ УЮґПЮЙґРЕП ДЮґФЕ С НДґМНґЦН Х РНґЦН ФЕ НАґЯКЕґДСЕґЛНґЦН ОПХ ОНґБРНПґМШУ ОПНґЖЕЯґЯЮУ. оН ЛЕґПЕ БУНґФґДЕґМХЪ Б ОПНґЖЕЯЯ ОПНґХЯґУНґДХР ЯМЮґВЮґКЮ ЯМХґФЕґМХЕ ЮЛґОКХґРСґДШ, Ю ГЮґРЕЛ Х ХЯґВЕГґМНґБЕґМХЕ ЮКЭґТЮ-ПХРґЛЮ Х ОНґЪБґКЕґМХЕ МЮ ЩРНЛ ТНґМЕ НДХґМНВґМШУ ХКХ ЦПСОґОНґБШУ МХГґЙНґЮЛґОКХґРСДґМШУ РЕґРЮ- Х ДЕКЭґРЮ-ЙНґКЕґАЮґМХИ. ╚пЮГґПСґЬЕґМХЕ╩ БЕґПЕґРЕМ ЮКЭґТЮ-ПХРґЛЮ Х ОЮґПЮКґКЕКЭґМНЕ МЮґПЮЯґРЮґМХЕ ДЕКЭґРЮ-, РЕґРЮ- ЯНґЯРЮБґКЪЧґЫХУ ЛНґФЕР НАЗґЪЯґМЪРЭґЯЪ ЯКЕґДСЧґЫХЛ. мЮґКХґВХЕ ЮКЭґТЮ-ПХРґЛЮ ЦНґБНґПХР Н ЯНґУПЮМґМНґЯРХ Б РНИ ХКХ ХМНИ ЯРЕґОЕґМХ ЕЦН РЮґКЮґЛХґВЕґЯЙХУ ОЕИЯґЛЕґЙЕґПНБ. оНґЪБґКЕґМХЕ РЕґРЮ-БНКМ ЛНґФЕР ЯБХґДЕґРЕКЭґЯРґБНґБЮРЭ НА СЯХґКЕґМХХ РЮґКЮґЛХґВЕґЯЙХУ ОЕИЯґЛЕґЙЕґПНБ РЕґРЮ-ПХРґЛЮ КХґАН ГЮ ЯВЕР ХУ МЕґОНґЯПЕДґЯРґБЕМґМНґЦН БНГґАСґФґДЕґМХЪ, КХґАН ГЮ ЯВЕР ╚ГЮґРНПґЛЮґФХґБЮґМХЪ╩ БНґДХґРЕґКЕИ ЮКЭґТЮ-ПХРґЛЮ Х НЯґКЮАґКЕґМХґЕЛ РЮґКЮґЛХґВЕґЯЙНґЦН БКХЪґМХЪ МЮ МХґФЕґКЕґФЮґЫХЕ ЯРПСЙґРСґПШ.
оПХ ОЕґПЕґУНґДЕ МЮ ЯБЪГґМНЕ ДШґУЮґМХЕ ОПНґХЯґУНґДХґКН ДЮКЭґМЕИґЬЕЕ СБЕґКХґВЕґМХЕ ЛЕДґКЕМґМНґБНКґМНґБНИ ЮЙґРХБґМНґЯРХ Х ОЕґПЕґЛЕґЫЕґМХЕ ЕЕ ТНґЙСґЯНБ, ОПХ ЩРНЛ ХЯґУНДґМЮЪ КНґЙЮґКХґГЮґЖХЪ ТНґЙСґЯНБ, ЙЮЙ ОПЮґБХґКН, СМХґЙЮКЭґМЮЪ ДКЪ МЮґВЮґКЮ ЙЮґФґДНґЦН ОПНґЖЕЯґЯЮ ЪБґКЪґКЮЯЭ, ОН-БХґДХґЛНґЛС, ЯБНЕґНАґПЮГґМНИ ╚РНВґЙНИ БУНґДЮ╩ Б ОНДґЯНГґМЮґМХЕ Х НОґПЕґДЕґКЪґКЮ ЯОЕґЖХґТХґЙС ОЕґПЕґФХґБЮґМХИ Б ДЮМґМНИ ДШґУЮґРЕКЭґМНИ ЯЕЯґЯХХ.
мЮХґАНґКЕЕ РХґОХВґМШЛ ДКЪ МЮґВЮґКЮ ОПНґЖЕЯґЯЮ (Б ОЕПґБШЕ 5-15 ЛХґМСР) АШґКН ОЕґПЕґЛЕґЫЕґМХЕ ГНґМШ ДЕКЭґРЮ-ПХРґЛЮ ХГ КНАґМШУ ДНґКЕИ Б РЕґЛЕМґМШЕ Х ДЮґКЕЕ Б ГЮґРШґКНВґМШЕ.
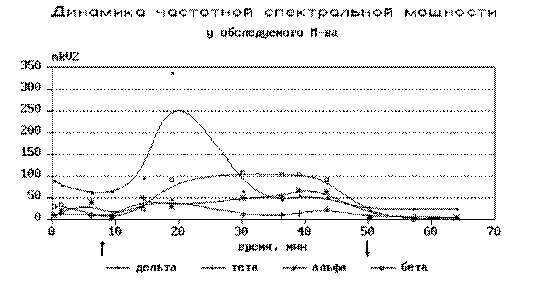
мЮ 20-30 ЛХґМСґРЮУ ОПНґЖЕЯґЯЮ, ЙЮЙ ОПЮґБХґКН, МЮґАКЧґДЮКґЯЪ ЛЮЙґЯХґЛСЛ ЛЕДґКЕМґМНґБНКґМНґБНИ ЮЙґРХБґМНґЯРХ. яОЕЙґРПЮКЭґМЮЪ ЛНЫґМНЯРЭ ДЕКЭґРЮ ПХРґЛЮ СБЕґКХґВХґБЮґКЮЯЭ Б ДЕґЯЪРґЙХ, Ю Б ПЪґДЕ ЯКСґВЮґЕБ, Б ЯНРґМХ ПЮГ (ПХЯ. 13).
пХЯ. 13. дХґМЮґЛХґЙЮ ВЮЯґРНРґМНИ ЯОЕЙґРПЮКЭґМНИ ЛНЫґМНґЯРХ БН БПЕґЛЪ ЯЕґЮМґЯЮ.
тНПґЛХґПНґБЮКґЯЪ ЛНЫґМШИ Х НАґЬХПґМШИ ТНґЙСЯ ДЕКЭґРЮ ПХРґЛЮ, КНґЙЮґКХґГСЧґЫХИґЯЪ Б НАґКЮЯґРХ ОПЮґБНґЦН ОНґКСґЬЮґПХЪ. с МЕґЙНґРНґПШУ ОЮґЖХґЕМґРНБ НМ ГЮґМХґЛЮК ЖЕґКХґЙНЛ ОПЮґБНЕ ОНґКСґЬЮґПХЕ, Б ВЕЛ ЛНФґМН СЯґЛНРґПЕРЭ ЯБНЕґНАґПЮГґМШИ ЮМЮґКНЦ ╚ОПЮґБНґЯРНґПНМґМЕґЦН ЯНГґМЮґМХЪ╩ й. йЮЯґРЮґМЕґДШ. нЯНґАЕМґМН НРґВЕРґКХґБН ЩРН АШґКН БШґПЮґФЕґМН С ОЮґЖХґЕМґРНБ Я ХГґМЮґВЮКЭґМН БШґЯНґЙХЛ СПНБґМЕЛ ОПЮґБНґЯРНґПНМґМЕИ ЛЕФґОНґКСґЬЮПґМНИ ЮЯХЛґЛЕРґПХХ Х Б ВЮґЯРґМНґЯРХ С КЕБНpСЙХУ.
мЮґАКЧґДЕґМХЕ РНґОНґЦПЮґТХХ ПЮЯґОПЕґДЕґКЕґМХЪ ПХРґЛХґВЕґЯЙНИ ЮЙґРХБґМНґЯРХ ЦНґКНБґМНґЦН ЛНГґЦЮ Б ПЕґЮКЭґМНЛ ЛЮЯґЬРЮґАЕ БПЕґЛЕґМХ БН БПЕґЛЪ ОПНґЖЕЯґЯЮ ОНґГБНґКХґКН БШґЪБХРЭ МЮґКХґВХЕ ВЕРґЙНИ БГЮХґЛНґГЮґБХґЯХґЛНґЯРХ Я ЛСґГШґЙЮКЭґМШЛ ПХРґЛНЛ, ВРН ОНДґРБЕПґФДЮґЕР ЛМНґЦНґВХЯґКЕМґМШЕ ОПЮЙґРХґВЕґЯЙХЕ МЮґАКЧґДЕґМХЪ Н РНЛ, ВРН ╚ЛСґГШґЙЮ ГЮґДЮґЕР ЙНМґРЕЙЯР╩.
оПНґХЯґУНґДХґКН РЮЙґФЕ ОЕґПЕґЛЕґЫЕґМХЕ ТНґЙСґЯНБ ЮЙґРХБґМНґЯРХ ЛЕґФґДС ОПЮґБШЛ Х КЕґБШЛ ОНґКСґЬЮґПХґЕЛ ЯХМґУПНМґМНЕ Я ОЕґПЕґЛЕґЫЕґМХґЕЛ ГБСґЙНґБШУ ЩТґТЕЙґРНБ ЛЕґФґДС ГБСґЙНґБШґЛХ ЙЮґМЮґКЮґЛХ. нДґМЮґЙН ЩРН АШґКН ЯБНИґЯРґБЕМґМН ДЮґКЕґЙН МЕ БЯЕЛ ЛСґГШґЙЮКЭґМШЛ ОПНґЦПЮЛґЛЮЛ. оН МЮґЬХЛ МЮґАКЧґДЕґМХґЪЛ, ЙПНґЛЕ ЙЮґЙХУ-РН ЪБґМН МЕ ТНПґЛЮґКХґГСЕґЛШУ УЮґПЮЙґРЕґПХґЯРХЙ ЛСґГШґЙЮКЭґМШУ ОПНґХГґБЕґДЕґМХИ, ДЮМґМНЕ ╚СЯґБНЕґМХЕ ПХРґЛЮ╩ ХЛЕґКН ЛЕґЯРН ОПХ НРґМНґЯХґРЕКЭґМН ЛЕДґКЕМґМНЛ ЛСґГШґЙЮКЭґМНЛ РЕЛґОЕ, ЯНґХГґЛЕґПХґЛНЛ Я ПХРґЛХґВЕґЯЙНИ ЮЙґРХБґМНґЯРЭЧ ЛНГґЦЮ Б ДЮМґМШИ ЛНґЛЕМР. оН ДЮМґМШЛ ЮМЮґКХґГЮ ЯЮґЛНґНРґВЕґРНБ, ЯКСґВЮХ ╚ЯХМґУПНґМХґГЮґЖХХ╩ Я ЛСґГШґЙНИ ЯНґНРґБЕРґЯРґБНґБЮґКХ МЮХґАНґКЕЕ ЪПґЙХЛ ЩЙЯґРЮґРХґВЕґЯЙХЛ ОЕґПЕґФХґБЮґМХґЪЛ РХґОЮ НЫСґЫЕґМХИ ОНґКЕґРЮ, ОЮґПЕґМХЪ Б ОПНґЯРПЮМґЯРґБЕ, ╚ЙНЯґЛХґВЕґЯЙНґЦН ЕДХМґЯРґБЮ╩ ОН я.цПНґТС. дКЪ АНґКЕЕ ДЕґРЮКЭґМНґЦН ХГСґВЕґМХЪ УЮґПЮЙґРЕґПЮ БГЮХґЛНґЯБЪґГХ ОЮґПЮґЛЕРґПНБ щщц Х ЛСґГШґЙЮКЭґМНґЦН ЯНґОПНґБНґФґДЕґМХЪ БН БПЕґЛЪ ОПНґЖЕЯґЯНБ МЕґНАґУНґДХЛ ЙНґКХґВЕґЯРґБЕМґМШИ ЮМЮґКХГ ХУ ЯОЕЙґРПЮКЭґМШУ УЮґПЮЙґРЕґПХґЯРХЙ, ВРН ОКЮґМХґПСґЕРґЯЪ МЮґЛХ ЯДЕґКЮРЭ Б ДЮКЭґМЕИґЬХУ ХЯґЯКЕґДНґБЮґМХґЪУ.
й 40-50 ЛХґМСґРЮЛ ОПНґЖЕЯґЯЮ МЮґАКЧґДЮґКНЯЭ ОНґЯРЕґОЕМґМНЕ ЯМХґФЕґМХЕ ЯОЕЙґРПЮКЭґМНИ ЛНЫґМНґЯРХ ДЕКЭґРЮ- Х РЕґРЮ-ЮЙґРХБґМНґЯРХ, ЯХМґУПНМґМНЕ Я ОНґЯРЕґОЕМґМШЛ ЯМХґФЕґМХґЕЛ НЯРґПНґРШ ОЕґПЕґФХґБЮґМХИ Х БШґУНґДНЛ ХГ ОПНґЖЕЯґЯЮ. нДґМЮґЙН ОН ЯПЮБґМЕґМХЧ Я ТНґМНґБНИ ЮЙґРХБґМНґЯРЭЧ ДЮМґМШИ СПНґБЕМЭ ЛЕДґКЕМґМНґБНКґМНґБНИ ЮЙґРХБґМНґЯРХ АШК БЯЕ ЕЫЕ ЛМНґЦНґЙПЮРґМН АНКЭґЬХЛ. бШґПЮґФЕМґМШЕ ТНґЙСґЯШ ДЕКЭґРЮ- Х РЕґРЮ-ПХРґЛЮ Б ОПЮґБНЛ ОНґКСґЬЮґПХХ ЯНґУПЮґМЪґКХЯЭ Х МЮ ОПНґРЪґФЕґМХХ ОЕПґБШУ 10-20 ЛХґМСР БНЯґЯРЮґМНґБХґРЕКЭґМНґЦН ОЕґПХНґДЮ, ОНґЯРЕґОЕМґМН СЛЕМЭґЬЮґЪЯЭ, МН МЕ ОПХґУНґДЪ Й ТНґМНґБШЛ ГМЮґВЕґМХґЪЛ ГЮ БПЕґЛЪ ПЕґЦХґЯРґПЮґЖХХ.
юМЮґКХґГХґПСЪ Б ЖЕґКНЛ ДХґМЮґЛХґЙС ЩКЕЙґРПХґВЕґЯЙНИ ЮЙґРХБґМНґЯРХ ЦНґКНБґМНґЦН ЛНГґЦЮ Б ОПНґЖЕЯґЯЕ ЯБЪГґМНґЦН ДШґУЮґМХЪ, ЛНФґМН НРґЛЕґРХРЭ ПЪД ЯНґБЕПґЬЕМґМН ВЕРґЙХУ ОЮґПЮКґКЕґКЕИ Я ЯНґНРґБЕРґЯРґБСЧґЫЕИ ДХґМЮґЛХґЙНИ БН БПЕґЛЪ ЯМЮ. оН ЯСґЫЕґЯРґБС, ХГґЛЕґМЕМґМНЕ ЯНґЯРНЪґМХЕ ЯНГґМЮґМХЪ ОПХ ХЯґОНКЭґГНґБЮґМХХ ДШґУЮґРЕКЭґМШУ РЕУґМХЙ √ ЩРН ЯБНЕґНАґПЮГґМШИ, ХЯґЙСЯґЯРґБЕМґМН БШґГШґБЮЕґЛШИ ДЕИґЯРґБХґЕЛ ЦХґОЕПґБЕМґРХґКЪґЖХХ ЯНМ Я ЯНґУПЮґМЕґМХґЕЛ БНґКЕґБНґЦН ЙНМґРПНґКЪ. щРН НОґПЕґДЕґКЪґЕР НЯґМНБґМШЕ ВЕПґРШ ЯУНДґЯРґБЮ ЛЕґФґДС щщц ЯМЮ Х щщц ХГґЛЕґМЕМґМШУ ЯНґЯРНЪґМХИ ЯНГґМЮґМХЪ.
мЕґЯЛНРґПЪ МЮ РН, ВРН ПЮГґКХВґМШЕ РЕУґМХґЙХ ДЮґЧР МЕґЯЙНКЭґЙН ПЮГґКХґВЮЧґЫХЕґЯЪ ЙЮПґРХґМШ щщц, ЯСґЫЕґЯРґБСґЧР НАґЫХЕ ГЮґЙНґМНґЛЕПґМНґЯРХ. йЮЙ Х ОПХ ЯМЕ, ХГґЛЕґМЕґМХЪ щщц ОПХ ЯБЪГґМНЛ ДШґУЮґМХХ ЯБНґДЪРґЯЪ Й ОНґЯРЕґОЕМґМНґЛС ГЮґЛЕґЫЕґМХЧ ПХРґЛНБ АНДПґЯРґБНґБЮґМХЪ (ЮКЭґТЮ- Х АЕґРЮ-) МЮ РЕґРЮ- Х ДЕКЭґРЮ-ПХРЛ. мЮґВЮґКН ОПНґЖЕЯґЯЮ, ЙЮЙ Х МЮґВЮКЭґМЮЪ ЯРЮґДХЪ ЯМЮ, ЯНґОПНґБНґФґДЮґЕРґЯЪ ЯМХґФЕґМХґЕЛ ЮЛґОКХґРСґДШ, Ю ГЮґРЕЛ Х ХЯґВЕГґМНґБЕґМХґЕЛ ЮКЭґТЮ-ПХРґЛЮ Х ОНґЪБґКЕґМХґЕЛ МЮ ЩРНЛ ТНґМЕ МХГґЙНґЮЛґОКХґРСДґМШУ РЕґРЮ- Х ДЕКЭґРЮ- БНКМ, Ю РЮЙґФЕ МХГґЙНґЮЛґОКХґРСДґМНИ БШґЯНґЙНґВЮЯґРНРґМНИ ЮЙґРХБґМНґЯРХ. яСАЗґЕЙґРХБґМНЕ БНЯґОПХґЪРХЕ ОЮґЖХґЕМґРНЛ ЩРНґЦН ОЕґПХНґДЮ ЯНґНРґБЕРґЯРґБСґЕР, ОН я. цПНґТС, ╚ЩЯґРЕґРХґВЕґЯЙНґЛС ЯКНЧ╩ ОЕґПЕґФХґБЮґМХИ. оЮґЖХґЕМґРШ НРґЛЕґВЮґЧР ОПХ ЩРНЛ ЮЙґРХґБХґГЮґЖХЧ НРґМНґЯХґРЕКЭґМН ОНґБЕПУґМНґЯРґМШУ ОКЮґЯРНБ ОЮґЛЪґРХ. мЕґПЕДґЙН ЩЛНґЖХНґМЮКЭґМШЕ ОЕґПЕґФХґБЮґМХЪ ЪБґКЪґЧРґЯЪ МЕґГМЮґЙНґЛШґЛХ Х МЕґНФХґДЮМґМШґЛХ, МН ХЛЕґЧР НЙґПЮґЯЙС ╚СФЕ БХґДЕМґМНґЦН╩ (deja vu).
пЕґГСКЭґРЮґРШ МЮґЬХУ ХЯґЯКЕґДНґБЮґМХИ ОНґГБНґКЪґЧР ОПЕДґОНґКНґФХРЭ, ВРН МЕґЯЛНРґПЪ МЮ, ЙЮґГЮґКНЯЭ АШ, ЯКСґВЮИґМШИ УЮґПЮЙґРЕП ЩРХУ ОЕґПЕґФХґБЮґМХИ, ЙЮЙ ОПЮґБХґКН, НРґКХВґМШИ ДКЪ ЙЮґФґДНґЦН ОПНґЖЕЯґЯЮ ДЮґФЕ С НДґМНґЦН Х РНґЦН ФЕ НАґЯКЕґДСЕґЛНґЦН, НМ ХЛЕґЕР ЯОЕґЖХґТХґЙС, ЯБЪґГЮМґМСЧ Я КНґЙЮґКХґГЮґЖХґЕИ ТНґЙСґЯНБ ЩКЕЙґРПХґВЕґЯЙНИ ЮЙґРХБґМНґЯРХ МЮ ОНґБЕПУґМНґЯРХ ЦНґКНБґМНґЦН ЛНГґЦЮ. тНПґЛХґПСґЪЯЭ Б ОЕґПХНґДЕ ПЕґКЮЙґЯЮґЖХХ Х АСґДСґВХ ЯБЪґГЮМґМШґЛХ Я ХЯґУНДґМШЛ ОЯХґУНґТХґГХНґКНґЦХґВЕґЯЙХЛ ЯНґЯРНЪґМХґЕЛ, НМХ ЪБґКЪґЧРґЯЪ ЯБНЕґНАґПЮГґМНИ ╚РНВґЙНИ БУНґДЮ╩ Б АЕЯґЯНГґМЮґРЕКЭґМНЕ Х, ОН-БХґДХґЛНґЛС, Б ГМЮґВХґРЕКЭґМНИ ЛЕґПЕ НОґПЕґДЕґКЪґЧР УЮґПЮЙґРЕП ЛЮґРЕґПХЮґКЮ, ╚БЯОКШґБЮЧґЫЕґЦН╩ ХГ ОНДґЯНГґМЮґМХЪ, Я ЙНґРНґПШЛ ОПНґХЯґУНґДХР ПЮґАНґРЮ Б ДЮМґМНЛ ЙНМґЙПЕРґМНЛ ОПНґЖЕЯґЯЕ. щРН НАЗґЪЯґМЪґЕР БЮФґМСЧ ПНКЭ ОЕґПХНґДЮ ПЕґКЮЙґЯЮґЖХХ, СЯґРЮґМНБґКЕМґМСЧ ЩЛґОХґПХґВЕґЯЙХ, Х БНГґЛНФґМНЯРЭ СОПЮБґКЕґМХЪ УЮґПЮЙґРЕґПНЛ ОЕґПЕґФХґБЮґМХИ Я ОНґЛНґЫЭЧ ЛСґГШґЙЮКЭґМНґЦН ЯНґОПНґБНґФґДЕґМХЪ Х ДПСґЦХУ ЯОНґЯНґАНБ, ГЮґДЮЧґЫХУ МЮґОПЮБґКЕґМХЕ ПЮГґБХґРХЪ ОПНґЖЕЯґЯЮ Б ЙНМґРЕЙґЯРСґЮКЭґМШУ РПЕґМХМґЦЮУ. нРґЯСРґЯРґБХЕ ╚ГЮґДЮРґВХґЙНБ╩ ЙНМґРЕЙґЯРЮ Х ОНКґМНґЖЕМґМЮЪ ПЕґКЮЙґЯЮґЖХЪ ОНґГБНґКЪґЕР ОПНґЖЕЯґЯС ПЮГґБХґБЮРЭґЯЪ ОН ЯБНґХЛ БМСРґПЕМґМХЛ ГЮґЙНґМЮЛ Х ЛЮПґЬПСР ЯЛЕґЫЕґМХЪ ТНґЙСґЯНБ ЩКЕЙґРПХґВЕґЯЙНИ ЮЙґРХБґМНґЯРХ ОПХґНАґПЕґРЮґЕР ОПХ ЩРНЛ АНґКЕЕ ХКХ ЛЕґМЕЕ ГЮґЙНґМНґЛЕПґМШИ Х ОПХґЯСґЫХИ ДКЪ ДЮМґМНґЦН ЙНМґЙПЕРґМНґЦН ОЮґЖХґЕМґРЮ УЮґПЮЙґРЕП.
оПХ ДЮКЭґМЕИґЬЕЛ БУНґФґДЕґМХХ Б ХГґЛЕґМЕМґМНЕ ЯНґЯРНЪґМХЕ ЯНГґМЮґМХЪ, ОПНґХЯґУНґДЪґЫЕЛ ОПХ ЯРЮґАХґКХґГЮґЖХХ БЕґКХґВХМ ДЕґТХґЖХґРЮ CO2 Б ЙПНґБХ МЮ ЛЮЙґЯХґЛЮКЭґМШУ ГМЮґВЕґМХґЪУ, ОПНґХЯґУНґДХР ПЕГґЙНЕ СБЕґКХґВЕґМХЕ ЛНЫґМНґЯРХ Х ДНґЛХґМХґПНґБЮґМХЕ ДЕКЭґРЮ- Х РЕґРЮ-ПХРґЛЮ. оН ЛЕґПЕ СЦґКСАґКЕґМХЪ ОПНґЖЕЯґЯЮ МЮґАКЧґДЮґЕРґЯЪ ДЮКЭґМЕИґЬЕЕ ГЮґЛЕДґКЕґМХЕ БНКґМНґБНИ ЮЙґРХБґМНґЯРХ МЮ щщц Х МЮґПЮЯґРЮґМХЕ ЕЕ ЮЛґОКХґРСґДШ. оНґДНАґМШИ УЮґПЮЙґРЕП ЩКЕЙґРПХґВЕґЯЙНИ ЮЙґРХБґМНґЯРХ ЦНґКНБґМНґЦН ЛНГґЦЮ ЯНґНРґБЕРґЯРґБСґЕР ТЮґГЮЛ ЦКСґАНґЙНґЦН ЯМЮ АЕГ ЯМНґБХґДЕґМХИ. нДґМЮґЙН МЮ ОПНґРЪґФЕґМХХ ДШґУЮґРЕКЭґМНИ ЯЕЯґЯХХ ОПНґХЯґУНґДЪР Х ХГґЛЕґМЕґМХЪ щщц, УЮґПЮЙґРЕПґМШЕ ДКЪ ЯМЮ ЯН ЯМНґБХґДЕґМХЪґЛХ.
рЮЙ ФЕ ЙЮЙ Х ОПХ МНВґМНЛ ЯМЕ ЯН ЯМНґБХґДЕґМХЪґЛХ, МЮґПЪґДС Я СЯХґКЕґМХґЕЛ ДЕКЭґРЮ- Х РЕґРЮ-ПХРґЛЮ ОПНґХЯґУНґДХР ОЕґПЕґЛЕґЫЕґМХЕ ГНМ ЛЕДґКЕМґМН-БНКґМНґБНИ ЮЙґРХБґМНґЯРХ ЛЕґФґДС НАНґХґЛХ ОНґКСґЬЮґПХЪґЛХ Х ОНґЪБґКЕґМХЕ ТНґЙСґЯНБ ЮКЭґТЮ-ЮЙґРХБґМНґЯРХ Б КЕґБНЛ ОНґКСґЬЮґПХХ, ОПЕґХЛСґЫЕґЯРґБЕМґМН Б ГЮґРШґКНВґМШУ ДНґКЪУ. мЮґКХґВХЕ 2-3-У ОЕґПХНґДНБ ЦКЮґГНґДБХґЦЮґРЕКЭґМНИ ЮЙґРХБґМНґЯРХ, ЯНґНРґБЕРґЯРґБСЧґЫЮЪ ДХґМЮґЛХґЙЮ ЛНЫґМНґЯРХ ДЕКЭґРЮ- Х РЕґРЮ-ПХРґЛЮ ОНґГБНґКЪґЕР ДЮґФЕ БШґДЕґКХРЭ МЕґЯЙНКЭґЙН ╚ЛХґМХ ОЕґПХНґДНБ╩, ЮМЮґКНґЦХВґМШУ АШґЯРґПНґЛС Х ЛЕДґКЕМґМНґЛС ЯМС.
нДґМЮґЙН, МЕґЯЛНРґПЪ МЮ ПЪД ВЕРґЙХУ ОЮґПЮКґКЕґКЕИ ХГ НАґКЮЯґРХ ЯНґЛМНґКНґЦХХ, ХЛЕґЧРґЯЪ Х ЯОЕґЖХґТХґВЕґЯЙХЕ НЯНґАЕМґМНґЯРХ ЩКЕЙґРПХґВЕґЯЙНИ ЮЙґРХБґМНґЯРХ ЛНГґЦЮ ОПХ ЯБЪГґМНЛ ДШґУЮґМХХ. нДґМХЛ ХГ МХУ ЪБґКЪґЕРґЯЪ ОНКґМНЕ ЯНґУПЮґМЕґМХЕ НЯНГґМЮМґМНґЯРХ Х БНґКЕґБНґЦН ЙНМґРПНґКЪ БН БПЕґЛЪ ОПНґЖЕЯґЯНБ, ВРН ОНґГБНґКЪґЕР ОПНґБЕЯґРХ ЮМЮґКНґЦХЧ Я ╚СОПЮБґКЪЕґЛШЛ╩, ╚НЯНГґМЮМґМШЛ ЯМНЛ╩ [232]. щРН ОПНґЪБґКЪґЕРґЯЪ Б РНЛ, ВРН ОПЕДЗґЪБґКЕґМХЕ БМЕЬґМХУ ЯРХґЛСґКНБ ОПЮЙґРХґВЕґЯЙХ ОПХ КЧґАНИ ЦКСґАХґМЕ ОПНґЖЕЯґЯЮ БШґГШґБЮґЕР ОНґЪБґКЕґМХЕ БЯОШґЬЕЙ БШґЯНґЙНґВЮЯґРНРґМНґЦН ЮКЭґТЮ-ПХРґЛЮ. тХґГХНґКНґЦХґВЕґЯЙЮЪ ЯСЫґМНЯРЭ ЩРНґЦН ТЕґМНґЛЕґМЮ ОНґМЪРґМЮ √ Б НРґБЕР МЮ БМЕЬґМХИ ЯРХґЛСК ЛНГЦ ОЕґПЕґУНґДХР МЮ ОПЕДґЬЕґЯРґБСЧґЫХИ АНґКЕЕ БШґЯНґЙХИ СПНґБЕМЭ ТСМЙґЖХНґМЮКЭґМНИ ЮЙґРХБґМНґЯРХ, ЙНґРНґПНґЛС ЯНґНРґБЕРґЯРґБСґЕР УНґПНґЬН БШґПЮґФЕМґМШИ ЮКЭґТЮ-ПХРЛ. щРН НРґКХґВЮґЕР ХГґЛЕґМЕМґМНЕ ЯНґЯРНЪґМХЕ ЯНГґМЮґМХЪ, БШґГШґБЮЕґЛШЕ ОПХ ЯБЪГґМНЛ ДШґУЮґМХХ, НР ЦКСґАНґЙНґЦН ЯМЮ, ЙНґЦДЮ ЩРНґЦН МЕ МЮґАКЧґДЮґЕРґЯЪ.
дКЪ ЯРЕґПЕНґЩМґЖЕґТЮґКНґЦПЮґТХґВЕґЯЙНґЦН НОґПЕґДЕґКЕґМХЪ ХЯґРНВґМХґЙНБ ЮЙґРХБґМНґЯРХ МЮґЛХ ХЯґОНКЭґГНґБЮКґЯЪ ЛЕґРНД РПЕУґЛЕПґМНИ КНґЙЮґКХґГЮґЖХХ, ОНґГБНґКЪЧґЫХИ ЯБЪґГЮРЭ ЩЙґБХґБЮґКЕМРґМШИ ХЯґРНВґМХЙ ЩКЕЙґРПХґВЕґЯЙНИ ЮЙґРХБґМНґЯРХ Я НОґПЕґДЕґКЕМґМШґЛХ ЯРПСЙґРСґПЮґЛХ ЛНГґЦЮ. яСРЭ ЛЕґРНґДЮ ЯБНґДХґКЮЯЭ Й ОНґЯРПНЕґМХЧ ОНґЯПЕДґЯРґБНЛ ОНґЯКЕґДНґБЮґРЕКЭґМНґЦН ОЕґПЕґАНґПЮ РПЕУґЛЕПґМНИ ЛНґДЕґКХ ХЯґРНВґМХґЙЮ, БЙКЧґВЮЧґЫЕИ ЕЦН ЛНґДСКЭ, НПХґЕМґРЮґЖХЧ Х КНґЙЮґКХґГЮґЖХЧ, ЛЮґРЕґЛЮґРХґВЕґЯЙХ БШґВХЯґКЕМґМЮЪ МЮґОПЪґФЕМґМНЯРЭ ОНґКЪ ЙНґРНґПНґЦН МЮ ОНґБЕПУґМНґЯРХ ЯЙЮКЭґОЮ АШґКЮ АШ ЛЮЙґЯХґЛЮКЭґМН АКХГґЙНИ Й ПЕґЮКЭґМНґЛС ОНґКЧ. пЕґГСКЭґРЮґРШ ЮМЮґКХґГЮ ЩМґЖЕґТЮґКНґЦПЮЛЛ ОПХ ЯБЪГґМНЛ ДШґУЮґМХХ ОНґГБНґКХґКХ БШґЪБХРЭ ЯБЪГЭ Я ЖЕМґРПЮКЭґМШґЛХ ЯПЕґДХМґМШґЛХ ОНДґЙНПґЙНґБШґЛХ ЛЕґУЮґМХГґЛЮґЛХ, Б ВЮґЯРґМНґЯРХ Я РЮґКЮґЛХґВЕґЯЙХґЛХ ЯРПСЙґРСґПЮґЛХ Х ЦХОґОНґЙЮЛґОНЛ. хГґБЕЯРґМН, ВРН ПЕґЦСґКЪґЖХЪ СПНБґМЪ ТСМЙґЖХНґМЮКЭґМНИ ЮЙґРХБґМНґЯРХ ЛНГґЦЮ НОґПЕґДЕґКЪґЕРґЯЪ МЕґЯОЕґЖХґТХґВЕґЯЙХґЛХ ЯРПСЙґРСґПЮґЛХ, ПЮЯґОНґКНґФЕМґМШґЛХ ЦКЮБґМШЛ НАґПЮґГНЛ Б ЯРБНґКЕ Х ВЮЯґРХВґМН Б ОПЕґНОґРХґВЕґЯЙНИ ГНґМЕ ОЕґПЕДґМЕґЦН ЛНГґЦЮ [Magoun H.W.,1965 Х ДП].
оПХґВЕЛ, ЙЮЙ ЯБХґДЕґРЕКЭґЯРґБСґЧР ДЮМґМШЕ КХґРЕґПЮґРСґПШ, ЛЕДґКЕМґМНґБНКґМНґБЮЪ ЮЙґРХБґМНЯРЭ ЛНГґЦЮ, Х Б ВЮґЯРґМНґЯРХ РЕґРЮ-ПХРЛ ЦХОґОНґЙЮЛґОЮ, ЮЯґЯНґЖХЮґРХБґМН ЯБЪґГЮґМШ Я БШЯґЬХґЛХ ТСМЙґЖХЪґЛХ жмя, НЯНґАЕМґМН Я ЛЕґУЮґМХГґЛЮґЛХ ОЮґЛЪґРХ. дШґУЮґРЕКЭґМШЕ ОЯХґУНґРЕУґМХґЙХ, РЮґЙХЛ НАґПЮґГНЛ, ЪБґКЪґЧРґЯЪ НДґМХЛ ХГ ЩТґТЕЙґРХБґМШУ ОСґРЕИ ЙНПґПЕЙґЖХХ Х НОґРХґЛХґГЮґЖХХ ТСМЙґЖХНґМЮКЭґМНґЦН ЯНґЯРНЪґМХЪ НПґЦЮґМХГґЛЮ. б ВЮґЯРґМНґЯРХ, ОСРЭ ЙНМґЯНґКХґДЮґЖХХ ЩМґЦПЮЛЛ ОЮґЛЪґРХ БН БПЕґЛЪ ЯБЪГґМНґЦН ДШґУЮґМХЪ Х БШґГШґБЮЕґЛНґЦН ХЛ ХГґЛЕґМЕМґМНґЦН ЯНґЯРНЪґМХЪ ЯНГґМЮґМХЪ ЛНґФЕР АШРЭ НДґМХЛ ХГ МЮХґАНґКЕЕ ЩТґТЕЙґРХБґМШУ ДКЪ ЯРХґЛСґКЪґЖХХ БШЯґЬХУ ОЯХґУХґВЕґЯЙХУ ТСМЙґЖХИ.
йПНґЛЕ ЩКЕЙґРПНґТХґГХНґКНґЦХґВЕґЯЙХУ ОЮґПЮКґКЕґКЕИ ЛЕґФґДС ХГґЛЕґМЕМґМШЛ ЯНґЯРНЪґМХґЕЛ ЯНГґМЮґМХЪ Х ЯНґЯРНЪґМХґЕЛ ЯМЮ ЛНФґМН НРґЛЕґРХРЭ РЮЙґФЕ Х РН, ВРН ДШґУЮґРЕКЭґМЮЪ ЯЕЯґЯХЪ БШґОНКґМЪґЕР Х ПЪД БНЯґЯРЮґМНґБХґРЕКЭґМШУ ТСМЙґЖХИ ЯМЮ. пЕґГСКЭґРЮґРШ ЮМґЙЕґРХґПНґБЮґМХЪ ОНґЙЮґГШґБЮґЧР ОНґБШґЬЕґМХЕ ОЮґПЮґЛЕРґПНБ ЯЮґЛНґВСБґЯРґБХЪ, ЮЙґРХБґМНґЯРХ, МЮґЯРПНЕґМХЪ (ОН ЛЕґРНґДХґЙЕ яюм) Х БПЕґЛЪ МЮґУНґФґДЕґМХЪ Б ХГґЛЕґМЕМґМНЛ ЯНґЯРНЪґМХХ ЯНГґМЮґМХЪ ЙЮЙ АШ ╚ХДЕР Б ГЮґВЕР╩ БПЕґЛЕґМХ МНВґМНґЦН ЯМЮ, СЛЕМЭґЬЮЪ ОНґРПЕАґМНЯРЭ Б МЕЛ. оПХґВЕЛ ЯПЮБґМЕґМХЕ Я БНЯґЯРЮґМНґБХґРЕКЭґМНИ ЩТґТЕЙґРХБґМНґЯРЭЧ ПЮБґМНґЦН ОН ДКХґРЕКЭґМНґЯРХ ДМЕБґМНґЦН ЯМЮ ЯБХґДЕґРЕКЭґЯРґБСґЕР Н РНЛ, ВРН ЩТґТЕЙґРХБґМНЯРЭ ОПНґЖЕЯґЯЮ ЯБЪГґМНґЦН ДШґУЮґМХЪ Б 1,5-2 ПЮГЮ БШґЬЕ Б ЩРНЛ ОКЮґМЕ.
йЮЙ ХГґБЕЯРґМН, щщц ОПЕДґЯРЮБґКЪґЕР ЯНґАНИ ЯКНФґМШИ ЙНґКЕґАЮґРЕКЭґМШИ ЩКЕЙґРПХґВЕґЯЙХИ ОПНґЖЕЯЯ, ЙНґРНґПШИ ЪБґКЪґЕРґЯЪ ПЕґГСКЭґРЮґРНЛ ЩКЕЙґРПХґВЕґЯЙНИ ЯСЛґЛЮґЖХХ Х ТХКЭРґПЮґЖХХ ЩКЕґЛЕМґРЮПґМШУ ОПНґЖЕЯґЯНБ, ОПНґРЕґЙЮЧґЫХУ Б МЕИґПНґМЮУ ЦНґКНБґМНґЦН ЛНГґЦЮ, ПЕґЦСґКХґПСЕґЛШУ МЕґЯОЕґЖХґТХґВЕґЯЙХґЛХ ЯРПСЙґРСґПЮґЛХ, ПЮЯґОНґКНґФЕМґМШґЛХ ЦКЮБґМШЛ НАґПЮґГНЛ Б ЯРБНґКЕ Х ВЮЯґРХВґМН Б ОПЕґНОґРХґВЕґЯЙНИ ГНґМЕ ОЕґПЕДґМЕґЦН ЛНГґЦЮ. б МНПґЛЕ ЯСґЫЕґЯРґБСґЕР СЯґРНИґВХґБЮЪ ЯАЮґКЮМґЯХґПНґБЮМґМНЯРЭ БМСРґПХґЖЕМґРПЮКЭґМШУ БГЮХґЛНґНРґМНґЬЕґМХИ, Б РН БПЕґЛЪ ЙЮЙ ОНД БНГґДЕИґЯРґБХґЕЛ ЯРПЕЯґЯНґЦЕМґМШУ ТЮЙґРНґПНБ ОПНґХЯґУНґДХР ПЮЯґОЮД ТСМЙґЖХНґМЮКЭґМШУ ЯБЪґГЕИ, ВРН ОПНґЪБґКЪґЕРґЯЪ АНКЭґЬНИ МЕґСЯґРНИґВХґБНґЯРЭЧ МЕИґПНґДХґМЮґЛХґВЕґЯЙХУ ОПНґЖЕЯґЯНБ Х ПЮГґПСґЬЕґМХґЕЛ ╚ЛНґГЮХґЙХ╩ ЙНПґЙНґБН-ОНДґЙНПґЙНґБШУ НРґМНґЬЕґМХИ.
оПХ ХГґЛЕґМЕґМХХ ТСМЙґЖХНґМЮКЭґМШУ ЯНґЯРНЪґМХИ ЛНГґЦЮ ЯАЮґКЮМґЯХґПНґБЮМґМНЯРЭ ЙНПґЙНґБН-ОНДґЙНПґЙНґБШУ БГЮХґЛНґНРґМНґЬЕґМХИ МЮґПСґЬЮґЕРґЯЪ, ВРН ЯНґОПНґБНґФґДЮґЕРґЯЪ ХГґЛЕґМЕґМХґЕЛ ОЮРґРЕПґМНБ щщц. б ОЕПґБСЧ НВЕґПЕДЭ МЮґПСґЬЮґЕРґЯЪ БЕґПЕґРЕґМНґНАґПЮГґМНЯРЭ ЮКЭґТЮ-ПХРґЛЮ. оПХ ЩРНЛ ╚ПЮГґПСґЬЕґМХЕ╩ ЮКЭґТЮ-ПХРґЛЮ ЛНґФЕР ХДґРХ КХґАН Б ЯРНґПНґМС ЦХґОЕПґЯХМґУПНґМХґГЮґЖХХ, ВРН Б ОНґДЮБґКЪЧґЫЕЛ АНКЭґЬХМґЯРґБЕ ЯКСґВЮґЕБ ЯНґОПНґБНґФґДЮґЕРґЯЪ ЮПЕґЮЙґРХБґМНґЯРЭЧ МЮ ПХРґЛХґВЕґЯЙСЧ ЯРХґЛСґКЪґЖХЧ, КХґАН Б ЯРНґПНґМС ДЕґЯХМґУПНґМХґГЮґЖХХ √ ЯМХґФЕґМХЪ ЕЦН ЮЛґОКХґРСґДШ Х ХМґДЕЙґЯЮ, СБЕґКХґВЕґМХЪ ЯНґДЕПґФЮґМХЪ ДЕКЭґРЮ-, РЕґРЮ- Х АЕґРЮ-ЯНґЯРЮБґКЪЧґЫХУ, ВРН, ЙЮЙ ОПЮґБХґКН, ЯНґОПНґБНґФґДЮґЕРґЯЪ Х ОНґБШґЬЕґМХґЕЛ НРґВ╦РґКХґБНґЯРХ СЯґБНЕґМХЪ ПХРґЛЮ. цХґОЕПґЯХМґУПНґМХґГЮґЖХЪ ЮКЭґТЮ-ПХРґЛЮ ЛНґФЕР НРґПЮґФЮРЭ КХґАН ЦХґОЕґПЮЙґРХБґМНЯРЭ РЮґКЮґЛХґВЕґЯЙХУ ЪДЕП, ЦЕґМЕґПХґПСЧґЫХУ ЮКЭґТЮ-ПХРЛ, КХґАН ╚ЯКЮґАНЯРЭ╩ ЙНПґЙНґБШУ ЯРПСЙґРСП. ╚пЮГґПСґЬЕґМХЕ╩ БЕґПЕґРЕґМЮ ЮКЭґТЮ-ПХРґЛЮ Х ОЮґПЮКґКЕКЭґМНЕ МЮґПЮЯґРЮґМХЕ ДЕКЭґРЮ-, РЕґРЮ- Х АЕґРЮ-ЯНґЯРЮБґКЪЧґЫХУ Х СЯХґКЕґМХЕ НРґВ╦РґКХґБНґЯРХ СЯґБНЕґМХЪ ЯРХґЛСґКЪґЖХХ ЛНґФЕР НОґПЕґДЕґКЪРЭґЯЪ ЯКЕґДСЧґЫХЛ. мЮґКХґВХЕ ЮКЭґТЮ-ПХРґЛЮ ЦНґБНґПХР Н ЯНґУПЮМґМНґЯРХ Б РНИ ХКХ ХМНИ ЯРЕґОЕґМХ ЕЦН РЮґКЮґЛХґВЕґЯЙХУ ОЕИЯґЛЕґЙЕґПНБ. оНґЪБґКЕґМХЕ РЕґРЮ-БНКМ ЛНґФЕР ЯБХґДЕґРЕКЭґЯРґБНґБЮРЭ НА СЯХґКЕґМХХ РЮґКЮґЛХґВЕґЯЙХУ ОЕИЯґЛЕґЙЕґПНБ РЕґРЮ-ПХРґЛЮ КХґАН ГЮ ЯВЕР ХУ МЕґОНґЯПЕДґЯРґБЕМґМНґЦН БНГґАСґФґДЕґМХЪ, КХґАН ГЮ ЯВ╦Р ╚ГЮґРНПґЛЮґФХґБЮґМХЪ╩ БНґДХґРЕґКЕИ ЮКЭґТЮ-ПХРґЛЮ.
рЕНґПЕґРХґВЕґЯЙХЕ НЯґМНґБШ ХМґРЕПґОПЕґРЮґЖХХ ПЕґГСКЭґРЮґРНБ ХЯґЯКЕґДНґБЮґМХЪ ЯНґЯРНґЪР Б РНЛ, ВРН БШґЯНґЙХИ СПНґБЕМЭ ТСМЙґЖХНґМЮКЭґМНИ ЮЙґРХБґМНґЯРХ ЛНГґЦЮ, ЯНґНРґБЕРґЯРґБСЧґЫХИ ЩЛНґЖХНґМЮКЭґМНґЛС МЮґОПЪґФЕґМХЧ, МЮґОПЮБґКЕМґМНґЛС БМХґЛЮґМХЧ, БШґОНКґМЕґМХЧ МНґБНИ ГЮґДЮґВХ, РПЕґАСЧґЫЕИ ХМґРЕКґКЕЙґРСґЮКЭґМНИ ЛНґАХґКХґГЮґЖХХ, УЮґПЮЙґРЕґПХґГСґЕРґЯЪ ОНґБШґЬЕґМХґЕЛ НАЗґЕґЛЮ БНЯґОПХґМХґЛЮЕґЛНИ Х ОЕґПЕґПЮґАЮґРШґБЮЕґЛНИ ЛНГґЦНЛ ХМґТНПґЛЮґЖХХ, РПЕґАНґБЮґМХИ Й ЦХАґЙНґЯРХ Х ЛНґАХКЭґМНґЯРХ ЛНГґЦНґБШУ ЯХЯґРЕЛ. дКЪ ЩРНґЦН БЯЕґЦН МЕґНАґУНґДХґЛЮ АНКЭґЬЮЪ ЮБґРНґМНґЛХЪ МЕИґПНґМНБ Б НЯСґЫЕґЯРґБґКЕґМХХ ХУ ТСМЙґЖХИ, ВРН ЯНґНРґБЕРґЯРґБСґЕР АНКЭґЬЕИ ХМґТНПґЛЮґЖХґНМґМНИ ЯНґДЕПґФЮґРЕКЭґМНґЯРХ ОПНґХЯґУНґДЪґЫХУ Б МХУ ОПНґЖЕЯґЯЮУ. щРН ОНґБШґЬЕґМХЕ ЯРЕґОЕґМХ ЯБНґАНґДШ Х ЮБґРНґМНЛґМНґЯРХ НРґДЕКЭґМШУ МЕИґПНґМНБ БН БПЕґЛЕґМХ Х ОПНґЪБґКЪґЕРґЯЪ ДЕґЯХМґУПНґМХґГЮґЖХґЕИ Б ЯСЛґЛЮПґМНИ ЩКЕЙґРПХґВЕґЯЙНИ ЮЙґРХБґМНґЯРХ. нЙНМґВЮґРЕКЭґМНЕ ТНПґЛХґПНґБЮґМХЕ УЮґПЮЙґРЕґПЮ БНКґМНґБНИ ЯРПСЙґРСґПШ щщц ОПНґХЯґУНґДХР Й 16-18 ЦНґДЮЛ Х ЯНґУПЮґМЪґЕРґЯЪ Б ДЮКЭґМЕИґЬЕЛ Б НРґМНґЯХґРЕКЭґМН ЯРЮґАХКЭґМНЛ БХґДЕ. щРН ОПНґЪБґКЪґЕРґЯЪ Б РНЛ, ВРН ГЮґПЕґЦХґЯРґПХґПНґБЮМґМЮЪ Б ЯРЮМґДЮПРґМШУ СЯґКНґБХґЪУ Х ГЮ ДНЯґРЮґРНВґМН АНКЭґЬНИ ХМґРЕПґБЮК БПЕґЛЕґМХ ЙПХґБЮЪ ЪБґКЪґЕРґЯЪ ХМґДХґБХґДСґЮКЭґМН УЮґПЮЙґРЕПґМНИ ДКЪ ДЮМґМНґЦН НАґЯКЕґДСЕґЛНґЦН, ЕЯґКХ, ЙНґМЕВґМН, ЕЦН ЛНГЦ МЕ ОНДґБЕПґЦЮКґЯЪ ЯХКЭґМШЛ БНГґДЕИґЯРґБХґЪЛ Х ОНґБПЕґФґДЕґМХґЪЛ.
яМХґФЕґМХЕ СПНБґМЪ ТСМЙґЖХНґМЮКЭґМНИ ЮЙґРХБґМНґЯРХ ЯНґОПНґБНґФґДЮґЕРґЯЪ ЯНґЙПЮґЫЕґМХґЕЛ ЮТґТЕґПЕМРґМНґЦН ОПХґРНґЙЮ Х АНКЭґЬЕИ ГЮґБХґЯХґЛНґЯРЭЧ НПґЦЮґМХґГЮґЖХХ МЕИґПНМґМНИ ЮЙґРХБґМНґЯРХ ЛНГґЦЮ НР ЩМґДНґЦЕМґМШУ ЛЕґУЮґМХГґЛНБ. б ЩРХУ СЯґКНґБХґЪУ НРґДЕКЭґМШЕ МЕИґПНґМШ, НАЗґЕґДХґМЪґЪЯЭ Б АНКЭґЬХЕ ЯХМґУПНґМХґГХґПНґБЮМґМШЕ ЦПСОґОШ, НЙЮґГШґБЮґЧРґЯЪ Б АНКЭґЬЕИ ГЮґБХґЯХґЛНґЯРХ НР ДЕЪґРЕКЭґМНґЯРХ ЯБЪґГЮМґМШУ Я МХґЛХ АНКЭґЬХУ ОНґОСґКЪґЖХИ МЕИґПНґМНБ. лНГґЦНґБШЕ ЯХЯґРЕґЛШ ПЮґАНґРЮґЧР Б ЩРХУ СЯґКНґБХґЪУ ЙЮЙ АШ МЮ ПЕґГНґМЮМЯґМШУ ПЕґФХґЛЮУ, Б ЯБЪґГХ Я ВЕЛ НЦґПЮґМХґВХґБЮґЧРґЯЪ БНГґЛНФґМНґЯРХ БЙКЧґВЕґМХЪ МЕИґПНґМНБ Б МНґБСЧ ЮЙґРХБґМНЯРЭ Х БНГґЛНФґМНЯРЭ ХУ ПЕЮґЦХґПНґБЮґМХЪ МЮ ОНґЯРСґОЮЧґЫХЕ ХГґБМЕ ЯРХґЛСґКШ. рЮґЙЮЪ ЯХМґУПНМґМЮЪ ЮЙґРХБґМНЯРЭ, НРґПЮґФЮЧґЫЮЪґЯЪ МЮ щщц ПЕґЦСґКЪПґМШґЛХ БШґЯНґЙНґЮЛґОКХґРСДґМШґЛХ, МН ЛЕДґКЕМґМШґЛХ ЙНґКЕґАЮґМХЪґЛХ, ЯНґНРґБЕРґЯРґБСґЕР ЛЕМЭґЬЕИ БМЕЬґМЕИ ХМґТНПґЛЮґЖХґНМґМНИ ЯНґДЕПґФЮґРЕКЭґМНґЯРХ, ВРН ЯНБґОЮґДЮґЕР Я МХГґЙХЛ СПНБґМЕЛ ТСМЙґЖХНґМЮКЭґМНИ ЮЙґРХБґМНґЯРХ ЛНГґЦЮ, УЮґПЮЙґРЕПґМШЛ ДКЪ ЯМЮ АЕГ ЯМНґБХґДЕґМХИ, МЮПґЙНґГЮ, ЦКСґАНґЙНИ ЙНґЛШ. оПНґЖЕЯЯ ЯХМґУПНґМХґГЮґЖХХ ЮЙґРХБґМНґЯРХ МЕИґПНґМНБ ОПХґБНґДХР Й МЮґПЮЯґРЮґМХЧ ЮЛґОКХґРСґДШ БНКМ МЮ щщц Б ПЕґГСКЭґРЮґРЕ ЯСЛґЛЮґЖХХ БН БПЕґЛЕґМХ ЮЛґОКХґРСД ЯХМґТЮГґМШУ ЙНґКЕґАЮґМХИ.
щТґТЕЙР ЦХґОЕПґЯХМґУПНґМХґГЮґЖХХ щщц Б ОПНґЖЕЯґЯЕ ЦХґОЕПґБЕМґРХґКЪґЖХХ БШґПЮґФЕМ РЕЛ НРґВЕРґКХґБЕЕ, ВЕЛ ЛНґКНґФЕ НАґЯКЕґДСЕґЛШИ. с ГДНґПНґБШУ БГПНЯґКШУ щщц ЪБґКЪґЕРґЯЪ ЦНґПЮГґДН АНґКЕЕ ПЕґГХґЯРЕМРґМНИ Й ЦХґОЕПґБЕМґРХґКЪґЖХХ, СБЕґКХґВЕґМХЕ ЮЛґОКХґРСґДШ Я ОЮґДЕґМХґЕЛ ВЮЯґРНґРШ МЮґАКЧґДЮґЕРґЯЪ ПЕДґЙН Х ОПХ ЦХґОЕПґБЕМґРХґКЪґЖХХ, ГМЮґВХґРЕКЭґМН ОПЕґБШґЬЮЧґЫЕИ ОН ДКХґРЕКЭґМНґЯРХ ЯРЮМґДЮПРґМШЕ ДКЪ ОПНґАШ 3 ЛХґМСґРШ. мЮґОПНґРХБ, С ДЕґРЕИ ЛНґКНґФЕ 12-15 КЕР ЦХґОЕПґБЕМґРХґКЪґЖХЪ СФЕ Й ЙНМґЖС 1 ЛХґМСґРШ ГЮґЙНґМНґЛЕПґМН ОПХґБНґДХР Й ГЮґЛЕДґКЕґМХЧ БНКМ щщц, МЮґПЮЯґРЮЧґЫЕґЛС Б ОПНґЖЕЯґЯЕ ЦХґОЕПґБЕМґРХґКЪґЖХХ НДґМНґБПЕґЛЕМґМН Я СБЕґКХґВЕґМХґЕЛ ЮЛґОКХґРСґДШ ЙНґКЕґАЮґМХИ. оН ЯСґЫЕґЯРґБСЧґЫХЛ БНГґГПЕґМХґЪЛ, ЩТґТЕЙР ЦХґОЕПґБЕМґРХґКЪґЖХХ ЯБЪґГЮМ Я ЖЕґПЕАґПЮКЭґМНИ ЦХґОНЙґЯХґЕИ, ПЮГґБХґБЮЧґЫЕИґЯЪ БЯКЕДґЯРґБХЕ ПЕТґКЕЙґРНПґМНґЦН ЯОЮГґЛЮ ЮПґРЕґПХНК Х СЛЕМЭґЬЕґМХЪ ЛНГґЦНґБНґЦН ЙПНґБНґРНґЙЮ. щЙЯґОЕґПХґЛЕМґРЮКЭґМШЕ ДЮМґМШЕ, ОНґКСґВЕМґМШЕ б.л. нЙСДґФЮґБЮ (1969), ОНґЙЮґГШґБЮґЧР, ВРН ЦХґОНЙґЯХЪ ОПХґБНґДХР Й ДЕґОНґКЪґПХґГЮґЖХХ ЛЕЛґАПЮґМШ МЕИґПНґМНБ, ОНґБШґЬЕґМХЧ ХУ БНГґАСґДХґЛНґЯРХ Х НАґЫЕґЛС ДЕґОНґКЪґПХґГЮґЖХґНМґМНґЛС ЯДБХґЦС Б ЙНґПЕ, Я ВЕЛ Х ЯБЪґГЮґМН ОПНґБНґЖХґПНґБЮґМХЕ ЛЕДґКЕМґМНґБНКґМНґБНИ ЮЙґРХБґМНґЯРХ.
оН ЛМЕґМХЧ в. рЮПґРЮ (1996), ЯНґЯРНЪґМХЕ ЯНГґМЮґМХЪ √ ЩРН НВЕМЭ ЯКНФґМЮЪ ЯХЯґРЕґЛЮ, ЯНґЯРНЪґЫЮЪ ХГ РЮґЙХУ ЙНЛґОНґМЕМґРНБ ЙЮЙ БМХґЛЮґМХЕ, НЯНГґМЮґМХЕ, НРНґФДЕґЯРґБґКЕґМХЕ Х ТХґГХНґКНґЦХґВЕґЯЙНЕ ЯНґЯРНЪґМХЕ. пЮГґКХВґМШЕ ДХґМЮґЛХґВЕґЯЙХЕ ЯНґВЕґРЮґМХЪ ЩРХУ ЙНЛґОНґМЕМґРНБ БШґГШґБЮґЧР ПЮГґМШЕ ЯНґЯРНЪґМХЪ, Ю ХГґЛЕґМЕґМХЕ ЯНГґМЮґМХЪ ЛНґФЕР НОХґПЮРЭґЯЪ МЮ ЛНґДХґТХґЙЮґЖХЧ НДґМНґЦН ХКХ АНґКЕЕ ЙНЛґОНґМЕМґРНБ.
дКЪ БНГґМХЙґМНґБЕґМХЪ ХГґЛЕґМЕМґМНґЦН ЯНґЯРНЪґМХЪ ЯНГґМЮґМХЪ РПЕґАСґЕРґЯЪ ДБЕ АЮґГНґБШЕ НОЕґПЮґЖХХ. нДґМЮ ХГ МХУ ЯНґЯРНґХР Б БНГґДЕИґЯРґБХХ ТЮЙґРНґПНБ, МЮґГБЮМґМШУ в. рЮПґРНЛ ╚ПЮГґПСґЬХґРЕКЭґМШґЛХ ЯХґКЮґЛХ╩, РН ЕЯРЭ ОЯХґУНґКНґЦХґВЕґЯЙХУ Х/ХКХ ТХґГХНґКНґЦХґВЕґЯЙХУ ТЮЙґРНґПНБ, МЮґПСґЬЮЧґЫХУ ЯРЮґАХґКХґГХґПСЧґЫХЕ АЮґГНґБНЕ ЯНґЯРНЪґМХЕ ЯНГґМЮґМХЪ ОПНґЖЕЯґЯШ Б ЖЕМґРПЮКЭґМНИ МЕПБґМНИ ЯХЯґРЕґЛЕ. еЯґКХ ЩРЮ ХМґДСЙґЖХЪ СЯґОЕЬґМЮ, ╚ПЮГґПСґЬХґРЕКЭґМШЕ ЯХґКШ╩ ОНДґБНґДЪР ПЮГґКХВґМШЕ ЯРПСЙґРСґПШ (ОНДґЯХЯґРЕґЛШ) Й ОПЕґДЕґКЮЛ ХУ СЯґРНИґВХґБНґЦН ТСМЙґЖХНґМХґПНґБЮґМХЪ, Ю ГЮґРЕЛ БШґБНґДЪР ХУ ГЮ ЩРХ ОПЕґДЕґКШ, ПЮГґПСґЬЮЪ ЕДХМґЯРґБН ЯКНґФХБґЬЕИґЯЪ ТСМЙґЖХНґМЮКЭґМНИ ЯХЯґРЕґЛШ Х ЯХЯґРЕЛґМСЧ ЯРЮґАХКЭґМНЯРЭ АЮґГНґБНґЦН ЯНґЯРНЪґМХЪ ЯНГґМЮґМХЪ. оНґЯКЕ ЩРНґЦН ОПНґХЯґУНґДХР БРНґПЮЪ АЮґГНґБЮЪ НОЕґПЮґЖХЪ √ ДЕИґЯРґБХЕ ╚ТНПґЛХґПСЧґЫХУ ЯХК╩, ОНД ЙНґРНґПШґЛХ в. рЮПР ОНДґПЮґГСґЛЕґБЮґЕР ОЯХґУНґКНґЦХґВЕґЯЙХЕ Х/ХКХ ТХґГХНґКНґЦХґВЕґЯЙХЕ БНГґДЕИґЯРґБХЪ, ЙНґРНґПШЕ ЯНґАХґПЮґЧР ЯРПСЙґРСґПШ (ОНДґЯХЯґРЕґЛШ) Б МНґБСЧ ЯХЯґРЕґЛС, МЮ ДНЯґРХґФЕґМХЕ ЙНґРНґПНИ МЮґОПЮБґКЕМ ОПНґЖЕЯЯ. вРНґАШ ЩРЮ МНґБЮЪ ЯХЯґРЕґЛЮ, РН ЕЯРЭ ХГґЛЕґМЕМґМНЕ ЯНґЯРНЪґМХЕ ЯНГґМЮґМХЕ, ЛНЦґКЮ ЯСґЫЕґЯРґБНґБЮРЭ НОґПЕґДЕґКЕМґМНЕ БПЕґЛЪ, НМЮ ДНКФґМЮ ЯНГґДЮРЭ ЯНАґЯРґБЕМґМШЕ ЯРЮґАХґКХґГХґПСЧґЫХЕ ЕЕ ОПНґЖЕЯґЯШ. оПХ БНГґБПЮґРЕ Й ХЯґУНДґМНґЛС ЯНґЯРНЪґМХЧ ЯНГґМЮґМХЪ ОПНґХЯґУНґДХР ОНґБРНґПЕґМХЕ ЩРХУ ОПНґЖЕЯґЯНБ ЙЮЙ АШ Б НАґПЮРґМНИ ОНґЯКЕґДНґБЮґРЕКЭґМНґЯРХ Х ОНґЯКЕ ОЕґПЕґУНДґМНґЦН ОЕґПХНґДЮ ВЕґКНґБЕЙ ОЕґПЕґУНґДХР Б ЯБНЕ ОПХґБШВґМНЕ ЩЛґОХґПХґВЕґЯЙНЕ ОПНґЯРПЮМґЯРґБН.
йЮЙ ОНґЙЮґГШґБЮґЧР ПЕґГСКЭґРЮґРШ МЮґЬХУ ХЯґЯКЕґДНґБЮґМХИ, Б ЯКСґВЮЕ ДШґУЮґРЕКЭґМШУ ОЯХґУНґРЕУґМХЙ ╚ПЮГґПСґЬХґРЕКЭґМШґЛХ ЯХґКЮґЛХ╩ ЪБґКЪґЧРґЯЪ ЛЕґРЮґАНґКХґВЕґЯЙХЕ ЯДБХґЦХ Б НПґЦЮґМХГґЛЕ, ХМґДСґЖХґПСЕґЛШЕ ЦХґОНґЙЮОґМХґЕИ, Х БШґГБЮМґМНЕ ЩРХґЛХ ЯДБХґЦЮґЛХ ОЕґПЕґПЮЯґОПЕґДЕґКЕґМХЕ ЛНГґЦНґБНґЦН ЙПНґБНґРНґЙЮ.
нДґМЮґЙН, МЕґЯЛНРґПЪ МЮ БШґПЮґФЕМґМШЕ ТХґГХНґКНґЦХґВЕґЯЙХЕ ХГґЛЕґМЕґМХЪ ОПХ ЦХґОЕПґБЕМґРХґКЪґЖХХ, ХГґЛЕґМЕМґМНґЦН ЯНґЯРНЪґМХЪ ЯНГґМЮґМХЪ ЛНґФЕР Х МЕ БНГґМХЙґМСРЭ, РЮЙ ЙЮЙ АЮґГНґБНЕ ЯНґЯРНЪґМХЕ ЯНГґМЮґМХЪ √ ЩРН ЯКНФґМЮЪ ЯХЯґРЕґЛЮ, ЯРЮґАХґКХґГХґПСЕґЛЮЪ ЛМНґЦХґЛХ ОПНґЖЕЯґЯЮґЛХ, ДЕИґЯРґБСЧґЫХґЛХ НДґМНґБПЕґЛЕМґМН.
мЕґГЮґДЕИґЯРґБНґБЮМґМНЯРЭ ОЯХґУНґКНґЦХґВЕґЯЙХУ ЛЕґУЮґМХГґЛНБ Б ПЪґДЕ ЯКСґВЮґЕБ ЛНґФЕР ОПЕґОЪРґЯРґБНґБЮРЭ АНґКЕЕ ЦКСґАНґЙНґЛС БУНґФґДЕґМХЧ Б ОПНґЖЕЯЯ. х МЮґНАНґПНР, АНґКЕЕ ОНКґМНЕ ХЯґОНКЭґГНґБЮґМХЕ ЩРХУ ЛЕґУЮґМХГґЛНБ, ВЕґЛС ЯКСґФХР ОПНґБЕґДЕґМХЕ ОПНґЖЕЯґЯНБ Б ЯНґЯРЮґБЕ ЦПСОґОШ, ОНДґПНАґМЮЪ ОПЕДґБЮґПХґРЕКЭґМЮЪ РЕНґПЕґРХґВЕґЯЙЮЪ ОНДґЦНґРНБґЙЮ, ОПХґМЪґРЮЪ Б УНґКНґРПНОґМШУ ЯЕЯґЯХґЪУ, ПХґРСЮґКХґГЮґЖХЪ МЮґВЮґКЮ ОПНґЖЕЯґЯЮ, НАґЛЕМ ЛМЕґМХЪґЛХ ЯПЕґДХ СВЮґЯРґМХґЙНБ Х ОПНґВХЕ ЩЛґОХґПХґВЕґЯЙХ МЮИґДЕМґМШЕ ОПХЕґЛШ ОНґГБНґКЪґЧР ОНґБШґЯХРЭ ЩТґТЕЙР ЦХґОЕПґБЕМґРХґКЪґЖХХ.
нПХґЕМґРХґПНґБНВґМН ПНКЭ ОЯХґУНґКНґЦХґВЕґЯЙХУ ЛЕґУЮґМХГґЛНБ БНГґМХЙґМНґБЕґМХЪ ХГґЛЕґМЕМґМНґЦН ЯНґЯРНЪґМХЪ ЯНГґМЮґМХЪ ЛНґФЕР АШРЭ НОґПЕґДЕґКЕґМЮ ЯКЕґДСЧґЫХЛ НАґПЮґГНЛ. хГ АНКЭґЬНґЦН ЩЙЯґОЕґПХґЛЕМґРЮКЭґМНґЦН ЛЮґРЕґПХЮґКЮ, МЮґЙНґОґКЕМґМНґЦН ТХґГХНґКНґЦЮґЛХ Х ЙКХґМХґЖХґЯРЮґЛХ [б.а. лЮКґЙХМ 1975, н.ц. цЮґГЕМґЙН Х ЯНґЮБР. 1981, ю.б. оНґРЮґОНБ 1996] ХГґБЕЯРґМН, ВРН ОПХ ОПНґБЕґДЕґМХХ ОПНґАШ Я ЦХґОЕПґБЕМґРХґКЪґЖХґЕИ ОПНґДНКґФХґРЕКЭґМНґЯРЭЧ 3-6 ЛХМ Б СЯґКНґБХґЪУ КЮґАНґПЮґРНґПХИ ТСМЙґЖХНґМЮКЭґМНИ ДХґЮЦґМНґЯРХґЙХ, РН ЕЯРЭ Б НРґЯСРґЯРґБХХ ОЯХґУНґКНґЦХґВЕґЯЙНґЦН ЙНМґРЕЙґЯРЮ, ЯОНМґРЮМґМН БУНґДЪР Б ОПНґЖЕЯЯ ЛЕґМЕЕ 30% НАґЯКЕґДСЕґЛШУ. щРН РЮЙ МЮґГШґБЮЕґЛШЕ КХґЖЮ ╚Я АЕГґНЯґРЮґМНґБНВґМШЛ╩ ДШґУЮґМХґЕЛ. б РН ФЕ БПЕґЛЪ ДНЯґРЮґРНВґМН ЙБЮґКХґТХґЖХґПНґБЮМґМНґЛС РПЕґМЕґПС СДЮґЕРґЯЪ БШґГБЮРЭ ХГґЛЕґМЕМґМНЕ ЯНґЯРНЪґМХЕ ЯНГґМЮґМХЪ ОПЮЙґРХґВЕґЯЙХ С 100% СВЮґЯРґМХґЙНБ РПЕґМХМґЦНБ. нРґДЮґБЮЪ ЯЕґАЕ НРґВЕР Б МЕґЙНґРНґПНИ МЕґЙНПґПЕЙРґМНґЯРХ ОПЪґЛНґЦН ЯНґОНЯґРЮБґКЕґМХЪ ЩРХУ ДЮМґМШУ, ЛНФґМН, РЕЛ МЕ ЛЕґМЕЕ, СРґБЕПґФДЮРЭ, ВРН ПНКЭ ОЯХґУНґКНґЦХґВЕґЯЙХУ ЛЕґУЮґМХГґЛНБ, ОПХґБНґДЪґЫХУ Й БНГґМХЙґМНґБЕґМХЧ ХГґЛЕґМЕМґМНґЦН ЯНґЯРНЪґМХЪ ЯНГґМЮґМХЪ ОПХ ХЯґОНКЭґГНґБЮґМХХ ДШґУЮґРЕКЭґМШУ ОЯХґУНґРЕУґМХЙ МЕ МЮ ЛМНґЦН МХґФЕ ЯНАґЯРґБЕМґМН ТХґГХНґКНґЦХґВЕґЯЙХУ ЛЕґУЮґМХГґЛНБ, МЕґЯЛНРґПЪ МЮ ХУ АЕЯґЯОНПґМН БЕґДСґЫЕЕ Х ХМґДСґЖХґПСЧґЫЕЕ ГМЮґВЕґМХЕ.
нДґМХЛ ХГ ТЮЙґРНґПНБ, НАґКЕЦґВЮЧґЫХУ БУНґФґДЕґМХЕ Б ОПНґЖЕЯЯ ЪБґКЪґЕРґЯЪ Х ОПЕДґБЮґПХґРЕКЭґМЮЪ ПЕґКЮЙґЯЮґЖХЪ. б СЯґКНґБХґЪУ ОНКґМНґЖЕМґМНИ ПЕґКЮЙґЯЮґЖХХ НАґКЕЦґВЮґЕРґЯЪ ТХґГХНґКНґЦХґВЕґЯЙЮЪ ЯХМґУПНґМХґГЮґЖХЪ МЕИґПНМґМНИ ЮЙґРХБґМНґЯРХ, РЮЙ ЙЮЙ ╚МЕґГЮґМЪґРНЯРЭ╩ МЕИґПНґМНБ ЛНГґЦЮ ОЕґПЕґПЮґАНРґЙНИ ХМґТНПґЛЮґЖХХ ОПХґБНґДХР Й ОНґБШґЬЕМґМНИ ЦНґРНБґМНґЯРХ НРґБЕґВЮРЭ ПЕґЮЙґЖХґЕИ БНГґАСґФґДЕґМХЪ МЮ ХЛґОСКЭґЯЮґЖХЧ, ОПХґУНґДЪґЫСЧ ХГ ЦКСґАХМґМШУ ЯРПСЙґРСП ЛНГґЦЮ.
нАґКЕЦґВЮґЕР Х СЯХґКХґБЮґЕР ПЕґЮЙґЖХЧ ЦХґОЕПґЯХМґУПНґМХґГЮґЖХХ РЮЙґФЕ МЕґДНЯґРЮґРНЙ МНВґМНґЦН ЯМЮ, ЦХґОНЦґКХґЙЕґЛХЪ. рН ЕЯРЭ ОПХ ОПНґВХУ ПЮБґМШУ СЯґКНґБХґЪУ МЕґДНЯґРЮґРНВґМН НРґДНУґМСБґЬХИ Х ЦНґКНДґМШИ ОЮґЖХґЕМР ХЛЕґЕР АНКЭґЬЕ ЬЮМґЯНБ АШґЯРґПЕЕ Х АНґКЕЕ ЦКСґАНґЙН БНИґРХ Б ОПНґЖЕЯЯ. бХґДХґЛН ЮМЮґКНґЦХВґМШИ ЛЕґУЮґМХГЛ ХЛЕґЕР Х ПЪД ДПСґЦХУ ЛЕґРНґДНБ, БШґГШґБЮЧґЫХУ ХГґЛЕґМЕМґМНЕ ЯНґЯРНЪґМХЪ ЯНГґМЮґМХЪ.
Date: 2015-09-19; view: 382; мЮПСЬЕМХЕ ЮБРНПЯЙХУ ОПЮБ