Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Комиссуральные волокна
|
|
Мозолистое тело (corpus callosum) является самой большой комиссурой новой коры. Волокна проходят через эту комиссуру из одного полушария в другое и затем расходятся в разных направлениях, соединяя все гомотопические области коры за исключением первичной зрительной коры (поле 17) и проекций руки и ноги в соматосенсорной коре. Волокна мозолистой лучистости пересекают волокна лучистого венца (corona radiata) и других ассоциативных пучков. Поскольку мозолистое тело короче больших полушарий, волокна идущие через клюв, колено или валик мозолистого тела дугообразно изгибаются в направлении лобных и, соответственно, затылочных полюсов. Эти дугообразные пучки волокон называются передними (малыми) (forceps frontalis) и задними (большими) (forceps occipitalis) щипцами мозолистой лучистости (см. рис. 8.15в). Передняя комиссура (comissura anterior) является спайкой между обонятельным мозгом, височными долями и, в меньшей степени, надклювной (супраростральной) лобной корой. Гиппокампова комиссура или спайка свода (comissura fornicis), в основном, соединяет задние колонки свода (ножки свода).
Функциональная организация коры
До 1861 года кора большого мозга считалась функционально однородной и поливалентной. Будучи «седалищем души и разума», функции коры должны были быть нераздельны. Эта теория считалась общепринятой после того как Пьер Флорен, сравнительный анатом и физиолог из Парижского университета опубликовал ее в 1824 г., исходя из своих опытов на животных (в основном, на птицах).
В 1861 г. снова в Париже Пьер Поль Брока выступил на Антропологическом Обществе и продемонстрировал локализацию функций в коре головного мозга человека. Он обнаружил, что афазия у одного из его больных возникла вследствие разрушения задней трети левой третьей лобной извилины, позднее названной «извилиной Брока». В пользу локализациии различных функций в мозговой коре вскоре были получены многочисленные доказательства как клиницистами, так и физиологами.
Хьюлингс Джексон (1864) изучал больных с фокальной эпилепсией и объяснил причину возникновения припадков раздражением прецентральной коры. Карл Вернике (1874) впервые описал сенсорную афазию и связал ее с поражением задней части первой височной извилины. Он также убедительно показал доминантность одного из полушарий. В США Вейр Митчелл из Филадельфии (1860) отметил, что мышцы на одной стороне тела иннервируются корой противоположного полушария головного мозга. Его опыты были прекращены из-за Гражданской войны. Роберт Бартолов из Цинциннати, штат Огайо (1874) лечил больного с разрушенным вследствие злокачественного заболевания черепом. Он использовал представившуюся возможность для наложения на кору электродов и обнаружил, что фарадизация определенной области коры вызывала сокращения конечностей на противоположной стороне тела и ипсилатеральный поворот головы. После публикации этих находок филантропы и религиозные фанатики из Цинциннати заставили его покинуть город.
Фриц и Гитциг (1870) были первыми, кто выделили двигательную кору у собак путем электрической стимуляции и показали соматотопическое представительство в коре контрлатеральных движений. Эти опыты были продолжены другими выдающимися физиологами, такими как Ферриер (1876), Бивор и Горслей (1890), Грюнбаум (1901) и Шеррингтон (1906). Что касается сенсорного коркового представительства, фон Гудден (1870) показал, что удаление обох глаз у молодого животного приводило к задержке развития затылочных долей. Мунк (1979) установил, что собаки без затылочных долей являются слепыми и что повреждение только коры поверхности затылочной доли вызывает «seelenblindheit», называемую теперь зрительной агнозией. В 1879 и 1892 гг. Ферриер отмечал, что при раздражении некоторых отделов височных долей животные навостряют уши. Дюссе де Баренн (1916) для раздражения коры вместо электрического тока применял стрихнин. Накладывая его на ограниченные участки постцентральной извилины он обнаружил, что раздражение различных участков этой извилины заставляло животное расчесывать различные участки тела. Таким образом стало возможным также подразделить сенсорную постцентральную извилину на соматотопические единицы.
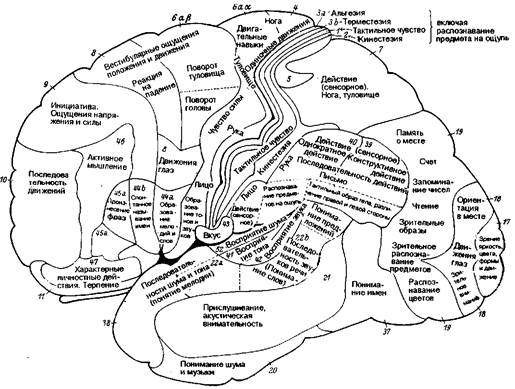
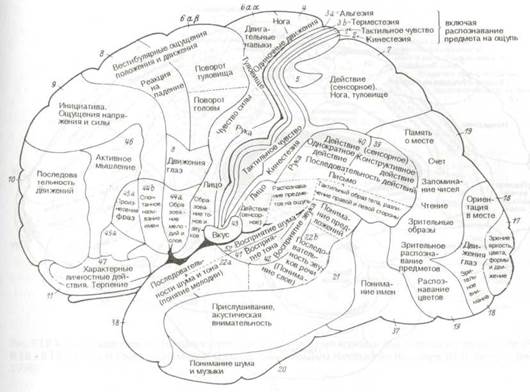
Рис. 8.16 Цитоархитектоническая локализация функций в коре большого мозга (по К. Клейсту). Конвекс левого полушария.
В этой попытке выделить и локализовать корковые функции истинный прогресс был медленным и полным противоречий. В качестве примера может быть приведена зрительная кора. Наши сегодняшние знания о ее функционировании и взимоотношении с сетчаткой и боковыми коленчатыми телами явились результатом драматических споров, продолжавшихся еще после 1-ой Мировой войны (Polyak, 1957; Lindenberg, 1977).
В это время (1934) ученик Вернике Карл Клейст опубликовал свою объемистую книгу «Патология мозга». Он анализировал клинические и патологоанатомические наблюдения над почти 300 солдатами, получившими черепно-мозговую травму во время 1-ой Мировой войны, и 106 госпитализированными больными с фокальными, в основном сосудистыми, мозговыми нарушениями с целью дальнейшего изучения функциональной организации головного мозга, включая локализацию основных психических функций.
Со времени смерти Вернике (1905) накопилось много полезной информации. Наиболее важным для Клейста казалось открытие, что серое вещество коры головного мозга являет собой совокупность множества отдельных единиц, каждая из которых имеет свою цито- миелоархитектонику (Campbell, 1905; E. Smith, 1907; Brodmann, 1909; O. Vogt, 1910; V. Economo и Koskinas, 1925; Beck, 1925; Rose, 1928). Он также заинтересовался тем обстоятельством, что образующие кору клеточные слои имеет различное функциональное значение (Nissl, 1908; Cajal, 1911; Kappers, 1920 и многие другие).
В конце концов, Клейст сделал вывод, что мозговая кора в целом состоит из набора архитектонических полей, представляющих преимущественно чувствительные сферы, и что каждая сфера состоит из истинно чувствительной зоны и двух более или менее распространенных зон — одной для двигательной и другой для психических функций. Он выделял следующие сферы: зрительную сферу в затылочной области, слуховую сферу в височной доле, осязательную сферу в центральной теменной области (включая двигательную область), вкусовую сферу в подцентральной оперкулярной области, лабиринтно-кинестезическую сферу в лобных извилинах, сферу внутренних чувств (область «я») в орбитальных и поясных извилинах и обонятельную сферу в грушевидной дольке и области гиппокампа.
Карта мозговой коры по Клейсту воспроизведена на рис. 8.16 и 8.17 как представляющая исторический интерес. Различные функции приписаны к обозначенным цифрами цитоархитектоническим полям по Бродману с некоторыми исправлениями по Экономо и Фогту. Сразу же после публикации этой карты Клейст был подвергнут суровой критике, особенно за привязку индивидуальных психических качеств к небольшим цитоархитектоническим полям. Привязка «я» (не путать с фрейдовским «эго») к очерченным корковым территориям была названа мозговой мифологией.
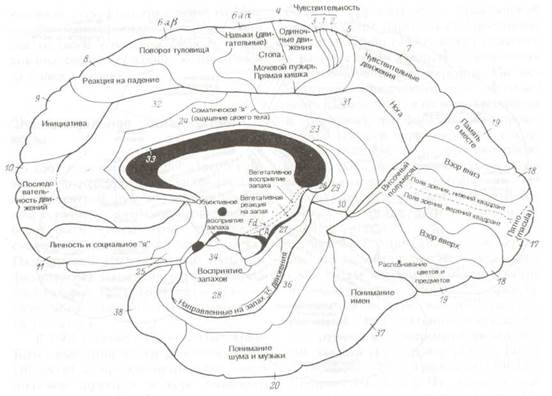
Рис. 8.17 Цитоархитектоническая локализация функций в коре большого мозга (по К.Клейсту). Медиальная поверхность правого полушария. (Рис. 8.16 и 8.17 взяты из: К. Kleist: Gehirnpathologie. In Handbuch der örztlichen Erfahrungen imWeltkrieg 1914/18, Bd. IV, Barth, Leipzig, 1922-1934).
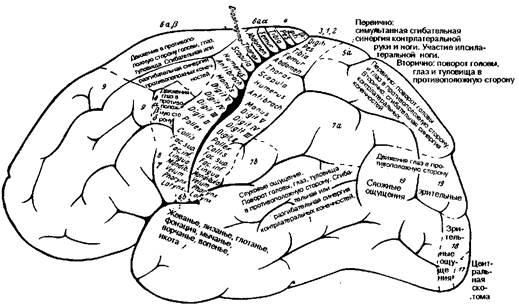
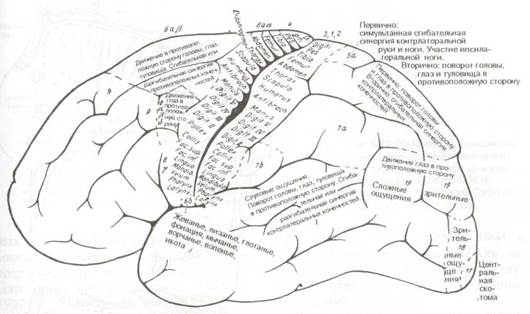
Рис. 8.18 Характер двигательных реакций в ответ на электростимуляцию корковых двигательных полей пациента. (Рис. 8.18 и 8.19 взяты из: 0. Foerster: Grosshirn. In: 0. Bumke, 0. Foerster (Eds), Handbuch der Neurologie, Bd.!V, Springer, Berlin, 1936).
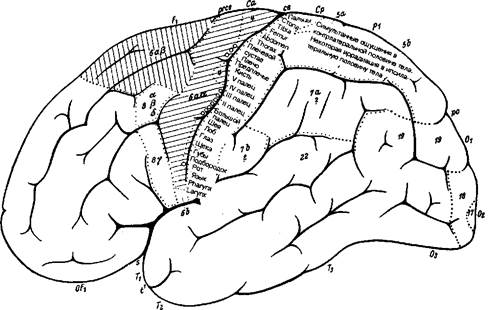
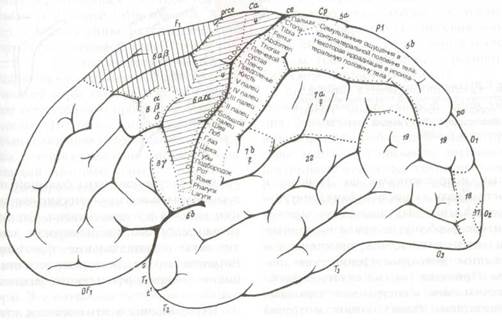
Рис. 8.19 Корковые зоны соместезии человека по Ферстеру.
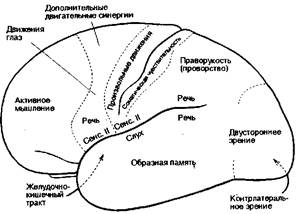
Рис. 8.21 Обобщенные функции корковых полей, определенные путем электростимуляции коры во время операции (из книги: Penfield W., Rasmussen Т.: The Cerebral Cortex of Man, Macmillan, N. Y.. 1950).
В последующие годы большой интерес вызнали психологические концепции холизма и гештальтизма. Существовало мнение о невозможности локализовать индивидуальные функциии. Впрочем, это не обескуражило клиницистов, нейроморфологов и физиологов.
Нет сомнения в возможности локализовать большое число функциональных нарушений. Эксперименты на животных по локализации функций нашли подтверждение при нейрохирургических операциях на человеке (Gushing, 1932; Foerster, 1936; Penfield, 1950 и другие). Новые данные могут быть получены путем электростимуляции коры головного мозга у оперируемых под местной анестезией больных (рис. 8.18, 8.19, 8.20 и 8.21). Дополнительные сведения удалось получить путем разрушения ограниченных участков коры у животных и людей (Минковский, 1917 и др.). К сожалению, раздражение или разрушение мозговой коры трудно отнести к физиологическим методам. Следовательно, вызванные электростимуляцией коры движения не соответствуют естественным произвольным движениям. Это было одним из побудительных мотивов продолжающегося поиска лучших методов исследования функций мозговой коры.
Электроэнцефалография принесла значительные успехи в этом направлении. Было обнаружено, что естественное или экспериментальное раздражение периферических рецепторов, таких как глаза или уши, вызывает регистрируемые изменения потенциалов мозговой коры. Эти вызванные потенциалы позволили довольно точно локализовать получающие стимулы области мозговой коры (Adrian, 1941; Woolsey, 1964 и др.). Вызванные потенциалы были записаны не только с коры, но и от глубинных структур мозга с помощью вживленных электродов.
В настоящее время можно с уверенностью утверждать что цитоархитектонические поля 3, 2 и 1 составляют первичную соматосенсорную область и что 17 поле получает зрительные импульсы, 41 поле — слуховые стимулы и 43 поле — вкусовые ощущения. Кроме того, установлено, что стимулы поступают в несколько первичных проекционных областей коры в строго определенном порядке (соматотопическом, ретинотопическом и тонотопическом).
Внедрение стереотактических методов намного продвинуло наши знания. Эти методы позволяют раздражать или разрушать любые глубинные структуры мозга и записывать вызванные потенциалы отдельных групп нейронов с помощью вживленных электродов.
Психохирургические операции, такие как лоботомия, лейкотомия и цингулотомия у больных с некоторыми психическими расстройствами (например, при шизофрении, неврозе навязчивых состояний и др.) позволили оценить значение передних лобных долей (префронтальной области) для некоторых поведенческих реакций.
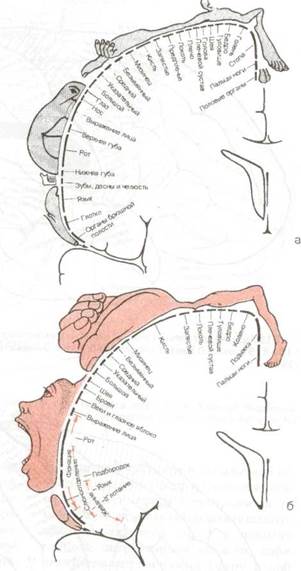
Рис. 8.20 Соматическое представительство в а - первичных сенсорных и б- первичных моторных корковых полях (из книги: Penfield W., Rasmussen Т.: The Cerebral Cortex of Man. Macmillan. N.Y., 1950).
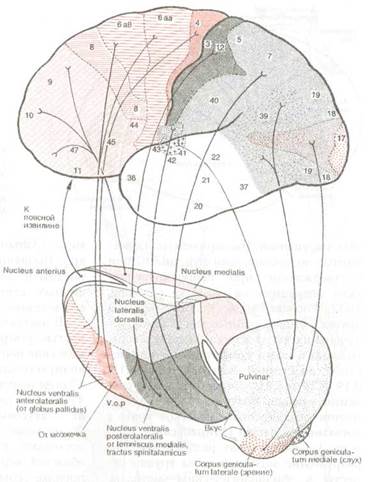
Рис. 8.22 Схема основных таламокортикальных связей и взаимоотношений индивидуальных корковых полей с таламическими ядрами.
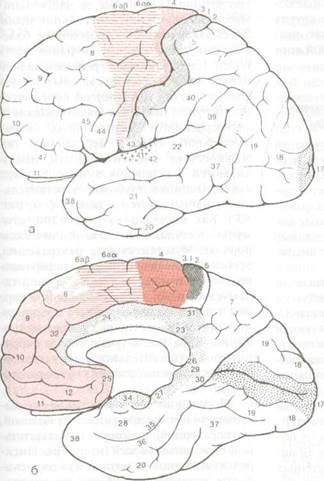
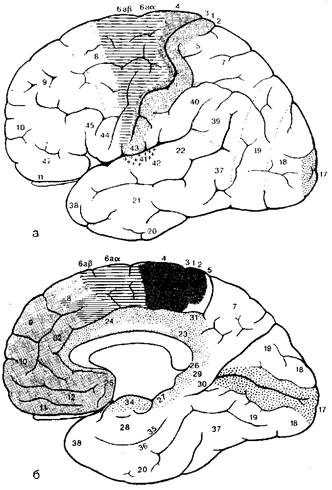
Рис. 8.23 Схема расположения первичных двигательных, чувствительных, зрительных вкусовых и слуховых полей, а также премоторных, префронтальных (включая орбитальные) и корковых лимбических зон: а - вид со стороны конвекса; б - вид с медиальной поверхности полушария.
В последние годы электронная микроскопия, флюоресцентная микроскопия и гистохимия принесли новое понимание структуры, метаболизма и функционирования нейронов. Очень важным оказалось введение микроэлектродов для записи потенциалов отдельных нервных волокон и нейронов. Этот метод, дополненный одной-двумя другими современными методиками, позволил физиологам картировать кору, головного мозга в зависимости от выполняемых ею функций. Вначале физиологи скептически.относились к большому числу полей, описанных как отдельные цитоархитектонические структуры Бродманом, Экономо и другими. Сегодня предполагаемое число полей по Хубелю и Визелю (1977) действительно лежит в пределах от 50 до 100.
Два вышеупомянутых автора являются признанными авторитетами в этом крайне запутанном и захватывающем вопросе. Они сосредоточились на зрительной коре (поле Бродмана 17) обезъян-макак. Полученные ими результаты могут быть суммированы в следующем виде: гранулярные клетки нижней трети четвертого слоя зрительной коры получают импульсы из бокового коленчатого тела и лучше всего реагируют на небольшие циркулярные стимулы, подобно ганглионарным клеткам сетчатки и нейронам коленчатого тела. Симметричные циркулярные поля информации в нижнем гранулярном слое должны быть, однако, перестроены, поскольку все клетки за пределами четвертого слоя лучше реагируют на специфически ориентированные сегменты линеарных стимулов. Клетки со схожей ориентацией линейных стимулов расположены в виде параллельных вертикальных колонок мозговой ткани, каждая из которых представляет собой «ориентационную колонку». Клетки, отвечающие на стимулы, исходящие только из одного глаза, занимают вертикальную колонку, прилежащую к другим клеточным колонкам, реагирующим на стимулы из другого глаза. Один набор таких «зрительно доминированных» колонок обслуживает оба глаза. По мнению авторов, сочетание одного набора колонок, обслуживающих все ориентации, и одного набора зрительно доминированных колонок является элементарной единицей первичной зрительной коры. Каждая такая вертикальная колонка занимает площадь 1 мм2 и имеет высоту 2 мм. Такое функцио  нальное разделение на вертикальные колонки было обнаружено также в других корковых полях. Впервые оно было описано в первичной соматосенсорной коре Маунткастлом в 1957г.
нальное разделение на вертикальные колонки было обнаружено также в других корковых полях. Впервые оно было описано в первичной соматосенсорной коре Маунткастлом в 1957г.
Date: 2015-07-02; view: 2662; Нарушение авторских прав