Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Внешнее строение
|
|
Конечный мозг или мозговая кора
Внешнее строение
Мозговая кора и ее белое вещество составляют самую большую часть конечного мозга. Полосатые тела, его небольшая часть, рассматривалась ранее.
Кора (cortex) выстилает наружную поверхность большого мозга и подобно плащу покрывает белое вещество (substantia alba) — отсюда и синонимы коры: мантия и плащ (pallium). В коре находятся тела нейронов, дендриты и немного аксонов, тогда как белое вещество состоит исключительно из миелинизированных аксонов. Внутренней поверхностью белого вещества являются стенки боковых желудочков.
Первоначально конечный мозг является частью круглого пузыря переднего мозга (конечный мозг + промежуточный мозг). На 4-й неделе эмбриональной жизни передний мозговой пузырь образует боковые карманы, которые впоследствие становятся полушариями большого мозга. К концу 4-го месяца эмбриональной жизни заканчивается развитие мозолистого тела в передне-заднем (ростро-каудальном) направлении. Разделенные межполушарной щелью полушария теперь соединены друг с другом с помощью спаек (комиссур): филогенетически более старая кора обонятельно-лимбической системы {архипаллиум + палеопаллиум, называемая также аллокортекс) — при помощи передней и еще меньшей гиппокамповой комиссур (commissura fonicis), и новая кора (неокортекс или не-опаллиум или изокортекс) — при помощи массивного мозолистого тела (corpus callosum). Размеры этой спайки столь велики потому, что новая кора намного обогнала в росте старую кору в процессе филогенетического развития мозга млекопитающего.
Для большей экономии места постоянно увеливающаяся кора образует все больше и больше складок в виде извилин, разделенных бороздами. В результате лишь треть общей площади коры человека находится снаружи, остальные две трети спрятаны в бороздах.
Особенности складок коры у разных млекопитающих очень различны, но для всех особей данного вида являются стереотипными и весьма характерными. Это справедливо и в отношении извилин человека. Их рисунок столь повторяем, что каждая большая извилина имеет свое имя (рис. от 8.4 до 8.8).
Некоторые борозды возникают в эмбриональном периоде раньше остальных и часто называются щелями. Самыми ранними являются коллаттеральная и носовая щели, отделяющие гиппокамповы извилины от других нижних височных извилин (образуются на третьем месяце внутриутробной жизни). Также рано формируются боковая или сильвиева щель (sulcus lateralis), центральная или роландова щель (sulcus centralis) и шпорная щель (sulcus calcari nus). Большинство основных извилин формируется после 6-го месяца внутриутробного развития. Развитие мелких извилин в результате разделения главных извилин неглубокими бороздками происходит не позднее раннего детства. Их рисунок строго индивидуален и различен в правом и левом полушарии одного и того же мозга.
Центральные щели на конвекситальной и медиальной поверхности полушария наиболее важны в структурном и функциональном отношении. Анатомически они являются границами между лобными (lobi frontales) и теменными (lobi parietales) долями. Функционально они разделяют переднюю соматомоторную область от задней соматосенсорной, о чем пойдет речь ниже. Кроме того, в каждом полушарии имеется еще две доли: затылочная (lobus occipitalis) и височная (lobus temporalis). Естественной границей затылочной доли является лишь затылочно-теменная щель (sulcus parietooccipitalis) на медиальной поверхности полушария и предзатылочная вырезка (incisura preoccipitalis) — маленькая зарубка на конце нижнего края височной доли. Прочие границы с теменной и височной долями достаточно условны (см. рис. 8.1, 8.2 и 8.3). Это довольно-таки символическое отражение сенсорного характера всех этих долей: затылочные доли служат зрению, а большая часть височных долей служит слуху. Таким образом, весь неопаллиум кзади от центральной щели является сенсорным: задняя центральная извилина, шпорная кора и извилины Гешля являются первичными мозговыми центрами телесных ощущений, зрения и слуха, соответственно. Вся кора между ними ответственна за высшую интегративную деятельность и взаимодействие между различными чувствами и их связи с другими частями мозга.
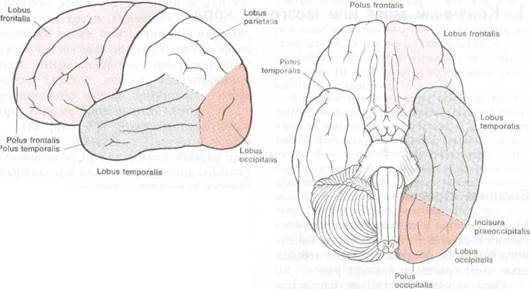
Рис. 8.1 Четыре доли большого мозга (вид со стороны свода левого полушария).
Рис. 8.3 Три доли большого мозга (вид с основания левого полушария после удаления левого мозжечка; орбитальная часть лобной доли часто называется орбитальной долей).

Рис. 8.2 Четыре доли большого мозга (вид с медиальной поверхности правого полушария).
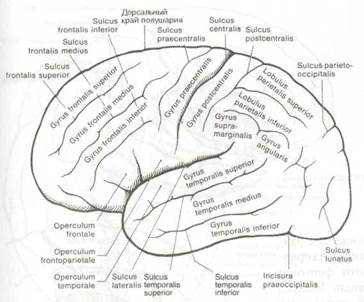
Рис. 8.4 Извилины и борозды большого мозга (вид сбоку).
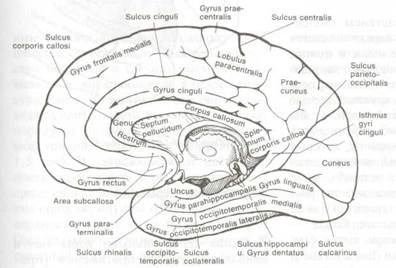
Рис. 8.5 Извилины и борозды большого мозга (медиальный вид).
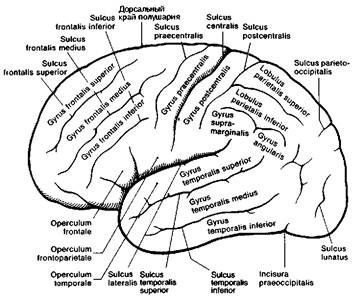
Рис. 8.4 Извилины и борозды большого мозга (вид сбоку).

Рис. 8.5 Извилины и борозды большого мозга (медиальный вид}.
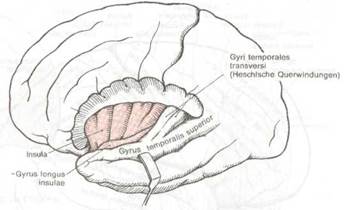
Рис. 8.6 Поперечные извилины Гешля и верх няя височная извилина.
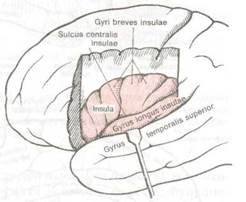
Рис. 8.7 Обнаженный островок (insula),
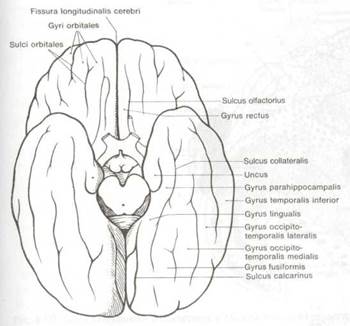
Рис. 8.8 Извилины и борозды на основании мозга.
По мере увеличения массы интегративной коры в процессе фило- и онтогенеза, височные доли сдвигаются кпереди. Это развитие объясняет почему некоторые волокна зрительной лучистости, исходящие из бокового коленчатого тела, идут кпереди прежде чем развернуться в противоположном направлении к шпорной коре. Этот феномен известен под названием петли Мейера, как было уже упомянуто ранее (см. гл. 3).
Более важным следствием смещения височных долей кпереди является возникновение боковой щели и полное закрытие островка (insula) височной доли. Глубоко спрятанные извилины островка ограничены полукружной щелью и называются также пятой долей. Чтобы увидеть их, необходимо резецировать лобные и теменные извилины, которые являются задней покрышкой (operculum) боковой щели и сдвинуть вниз височную долю как показано на рис. 8.6 и 8.7. При этом становятся также видны поперечные извилины Гешля — первичные корковые центры слуха (см. рис. 8.6) и порог островка (limen insulae), соединяющий островок с передней третью височной доли. Порог островка отношения к слуху не имеет.
Лимбическая доля Брока, о которой говорилось ранее (см. гл. 5) может считаться шестой долей конечного мозга.
Наконец, далее будет показано, что базальная часть лобной доли заслуживает выделения в самостоятельную седьмую долю конечного мозга — орбитальную долю.
Внутреннее строение
Кора
Толщина серой полоски коры почти одинакова на всем протяжении (приблизительно 4 мм). Она немного толще на вершине извилин чем в глубине борозд. Лишь небольшие участки коры имеют особое макроскопическое строение. Старая кора гиппокампа отличается завитком тонкой коры, образующим Аммонов рог. Самой толстой (приблизительно 5 мм в толщину) является двигательная кора передней центральной извилины. Она также менее отграничена от белого вещества по сравнению с остальной корой, поскольку здесь имеется относительно большое количество нейронов в так называемом седьмом слое, внедряющихся в белое вещество. Рядом расположена сенсорная кора задней центральной извилины, которая, наряду со шпорной корой, является самой тонкой (толщина ее не превышает 1,5 мм). В зависимости от насыщенности серого цвета коры на поперечном срезе иногда можно различить две тонкие полоски, идущие параллельно ее поверхности и друг другу по центру коры. Они называются линиями Бейларгера. В шпорной коре различима только одна полоска. Это линия Геннари — аналог наружной линии Бейларгера. Эти полоски белого цвета, потому что состоят из миелиновых волокон.

Рис, 8.8 Извилины и борозды на основании мозга.
Мозговая кора была разделена на множество полей в соответствии с различиями в архитектонике тканевых составляющих: нейронов, миелиновых волокон и кровеносных сосудов. Поэтому говорят о цитоархитектонике, миелоархитектонике и ангиоархитектонике. В последние годы благодаря гистохимическим исследованиям Фриде и других авторов заговорили о химиоархитектонике.
Сведениями по цитоархитектонике мы обязаны Бродману, Кемпбеллу, О. Фохту, фон Экономо, Коскинасу, фон Бонину и другим. Миелоархитектоника изучена благодаря Оскару и Цецилии Фохт и их ученикам. Пионером в изучении ангиоархитектоники был Р. А. Пфейфер.
Цитоархитектонические поля головного мозга человека по Бродману показаны на рис. 8.9. Следует подчеркнуть, что поля были пронумерованы в порядке их изучения. Никакого отношения к функциональным свойствам полей эта нумерация не имеет.
Архитектоника филогенетически старого аллокортекса мало дифференцирована по сравнению с новой корой, как видно на рис. 8.10, изображающем Аммонов рог гиппокампа и очень маленькую полоску зубчатой коры, состоящую из мелких плотно расположенных нейронов. Эта кора повторяет изгибы облитерированной гиппокамповой щели (sulcus hippocampi) и окружает концевую пластинку коры Аммонова рога, продолжающуюся в виде полукруга вокруг другой стороны гиппокамповой щели. Указанная кора состоит из других равновеликих пирамидных и двойных пирамидных клеток. Кора рога расширяется и продолжается в направлении подножья, или подставки (subiculum), кора которого местами состоит из четырех слоев. Второй слой состоит из множества островков нейронов, напоминающих аналогичные островки филогенетически старой обонятельной области. Кора подножья постепенно переходит в шестислойную кору (парагип-покампова извилина). Примерно на уровне коллатеральной щели кора представляет собой типичные шесть слоев изокортекса новой коры.
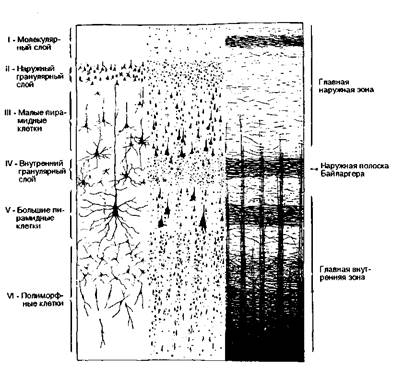
Архитектонику новой коры иллюстрирует рис. 8.11, где показана цитоар-хитектоника при импрегнации серебром и при окраске по Нисслю. Третья (правая) колонка демонстрирует богатый узор миелоархитектоники. Можно различить следующие слои нейронов начиная с поверхности коры.
Первый или молекулярный слой: содержит лишь небольшое число мелких нейронов (клетки Кахала). Дендриты этих клеток идут тангенциально внутри первого слоя, тогда как аксоны направляются к белому веществу. Эти клетки получают импульсы из пирамидных и веретенообразных клеток других корковых полей полушария, что объясняет большое число тангенциально идущих волокон.
Второй или наружный гранулярный слой: Нейроны его невелики и гранулярны. Между ними находится небольшое число мелких пирамидных клеток. Дендритные контакты происходят в пределах этого слоя.
Третий или наружный пирамидный слой: состоит из пирамидных клеток с широким основанием. Аксоны пирамидных клеток направляются к белому веществу и уже в этом слое являются миелинизированными. В белом веществе они идут как проекционные, ассоциативные или комиссуральные волокна. Отходящие от верхнего края клеток дендриты простираются в первый молекулярный слой. Остальные дендриты ветвятся в основном в наружном пирамидном слое.
Четвертый или внутренний гранулярный слой: аналогичен наружному гранулярному слою. Гранулярные клетки получают импульсы в основном по таламокортикальным путям. Подобно наружному гранулярному слою здесь содержатся многочисленные релейные станции с интернунциальными нейронами и системами обратной связи. В то время как волокна наружного пирамидного слоя в основном расположены радиально, волокна внутреннего гранулярного слоя, как правило, идут тангенциально, образуя наружную полоску Байларгера. Большинство этих волокон вероятно принадлежат нейронам особых таламических ядер.
Пятый или внутренний пирамидный слой: Пирамидные клетки этого слоя средних или больших размеров. В области прецентральной извилины в этом слое находятся гигантские пирамидные клетки (клетки Беца). Аксоны этих клеток окружены особенно толстыми миелиновыми оболочками. Они образуют корковоядерные и кортикоспинальные пути. Этот слой также содержит тангенциально ориентированные миелиновые волокна, образующие внутреннюю полоску Байларгера.
Шестой или мультиформноклеточный слой: клетки частично являются измененными пирамидными клетками, а частично — треугольными или веретенообразными клетками. Нейронов этого слоя меньше на границе с пятым слоем и больше на границе с белым веществом. Отходящие волокна соединяют этот слой с другими корковыми областями и с подкорковыми ядрами.
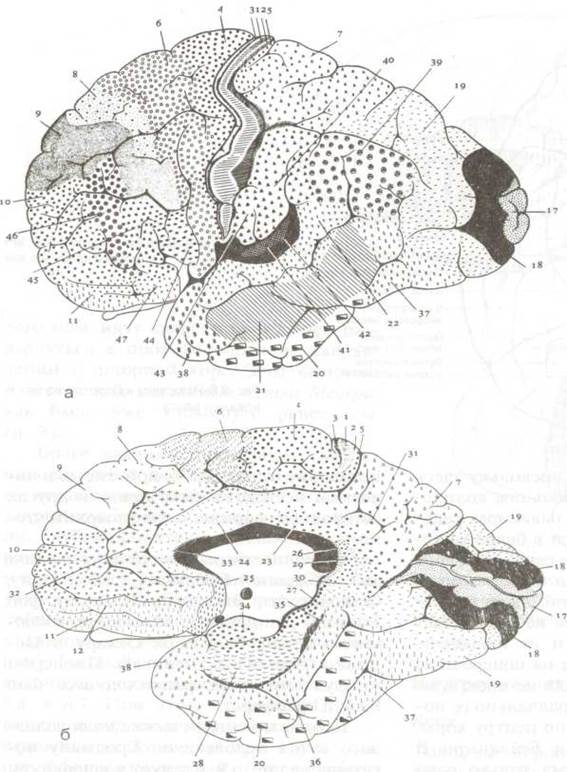
Рис. 8.9 Цитоархитектонические поля коры большого мозга: а конвекс левого полушария, б медиальная часть правого полушария. Цифрами обозначены корковые поля (по Бродману, из книги Bargmann, W. Histologie und mikroscopische Anatomie des Menschen, 6 Aufl. Thieme, Stuttgart, 1967.)
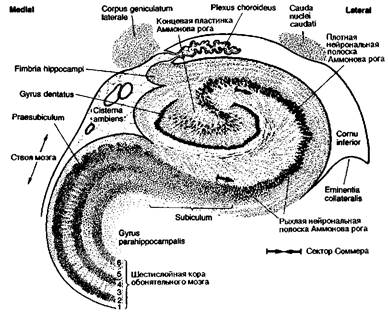
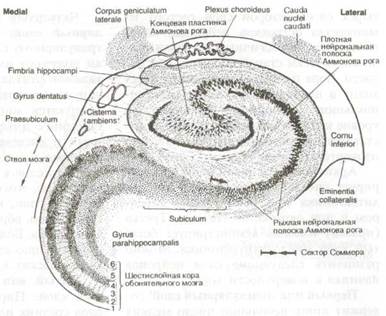
Рис. 8.10 Цитоархитектоника аллокортекса в области Аммонова рога. Гиппокамп в переводе с греческого означает «морской конек». При рассматривании рисунка справа парагиппокампова извилина образует шейку и завиток, напоминающий хвост морского конька. Эта её часть названа Аммоновым рогом в честь Аммона — древнеегипетского бога солнца, поскольку напоминает также бараний рог (священным животным Аммона считался баран и само божество часто изображалось в виде человека с бараньей головой).

Date: 2015-07-02; view: 3955; Нарушение авторских прав