Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Механизмы сокращения клеток сердечной мышцы
|
|
Сердечная мышца относится к возбудимым тканям, клетки которых в ответ на тот или иной раздражитель (электрический, химический, механический) могут генерировать электрические потенциалы действия – характерные изменения мембранного потенциала клетки, приводящие к ее возбуждению [1, 7]. Более того, некоторые виды клеток могут возбуждаться самопроизвольно. В основе механизма генерации потенциала действия клетками лежит изменение проницаемости мембран клеток для некоторых ионов (натрия, кальция, калия), активирующие ионные токи через специальные структуры клеточной мембраны — ионные каналы. Процессы, обеспечивающие сокращение клетки в ответ на электрическую стимуляцию объединяются понятием электромеханического сопряжения в сердечных клетках. Для мышечного сокращения необходимо повышение внутриклеточной концентрации Са2+ от уровня покоя (около 0.1 мкМ) до 1 мкМ [8].
Сердечные мышечные клетки (кардиомиоциты) включают в себя два основных типа по структуре и функциям: кардиомиоциты рабочего миокарда и клетки водителей сердечного ритма. Первый тип отвечает за механическое сокращение сердца, второй – за формирование устойчивого ритма сердечных сокращений [9].
Структура сердечной клетки является достаточно сложной [9, 10]: каждая клетка состоит из нескольких десятков повторяющихся сократительных единиц – саркомеров, длина которых около 2 мкм; поперечных трубочек (Т-трубочки, Т-тубулы), представляющих собой глубокие впячивания мембраны, опоясывают весь кардиомиоцит и локализуются у концов каждого саркомера. Сокращение саркомера обеспечивается мышечными волоконами (миофибриллами), расположенными вдоль саркомера и опоясанными сложной сетью внутриклеточных хранилищ ионов кальция, включающих продольный саркоплазматический ретикулум (СР), терминальные цистерны (ТЦ) (или люмен), примыкающие к мембране T-тубул (см. рис. 1.1б, рис. 1.3).
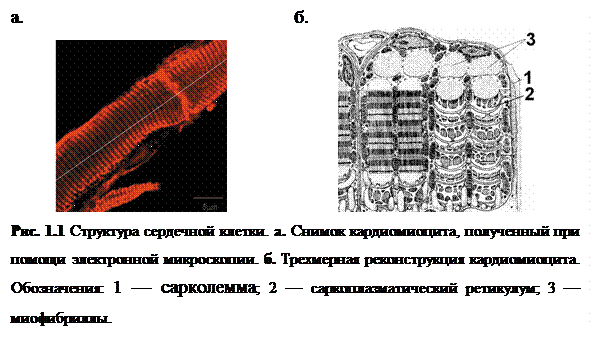
На мембране СР в местах контакта СР с Т-тубулами располагается группа специализированных Са2+-высвобождающих каналов, сопряженных с рианодиновыми рецептороами (RyR-каналы), через которые происходит высвобождение Са2+ из СР. Высвобождение носит триггерный характер, так как открытие RyR-каналов происходит в ответ на приток относительно небольшого количества Са2+ через ионные каналы, расположенных на мембране клетки – каналы L-типа или дигидропиридиновые рецепторы (см. рис.1.2, 1.3)[8, 10].

На мембране СР RyR-каналы образуют группы (кластеры), состоящие из нескольких сотен каналов. Существуют структуры, состоящие из кластера RyR-каналов вместе с несколькими L-каналами клеточной мембраны, люменом СР и диадным пространством между мембранами СР и примыкающими Т-тубулами. Данные структуры называются Са2+ высвобождающими единицами (Release Unit) (ВЕ, рис.1.2). В кардиомиоците насчитывается несколько десятков тысяч ВЕ; они формируют кальций-высвобождающую систему клетки (рис. 1.3).
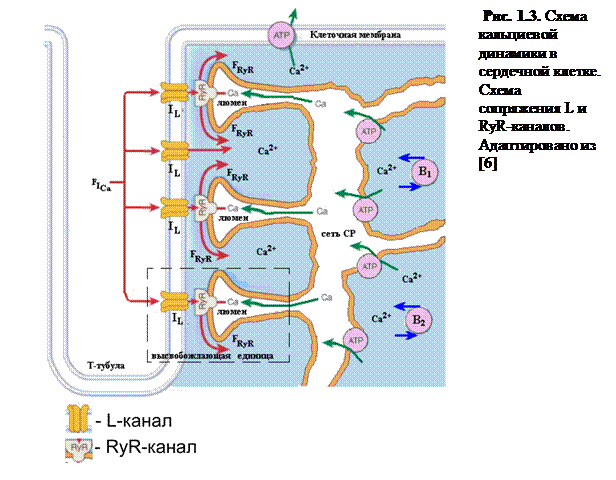
В связи с тем, что высвобождение Са2+ из СР осуществляется через группы RyR-каналов, исследование их динамических свойств является важной задачей в изучении процессов электромеханического сопряжения.
1.2 Рианодиновый рецептор – основной элемент управления кальциевой динамикой в клетке
RyR-канал является гигантским макромолекулярным комплексом (рис. 1.4), тетрамером, объединяющим четыре субъединицы (полипептидные цепи) с молекулярной массой 567 кДа каждая [2, 13]. Активность RyR модулируется под действием растительного алкалоида рианодина из коры Ryania speciosa, что и определило его название «рианодиновый рецептор». Существуют три изоформы RyR-каналов: RyR1 (клетки скелетной мускулатуры и мозжечка), RyR2 (клетки сердечной мышцы и мозга), RyR3 (клетки гладкой мускулатуры). Они различаются по своей структуре и динамическим параметрам.
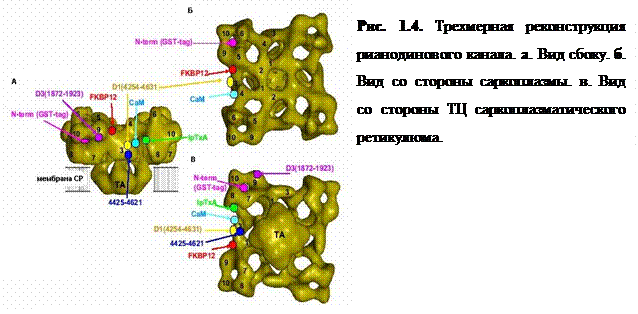 Как уже было сказано выше, RyR-каналы располагаются на мембране СР; при переходе в открытое состояние через них происходит высвобождение ионов Са2+ из просвета ТЦ (люмена). Согласно данным электронной микроскопии [12], RyR-канал имеет форму четырехлистника со стороной 27 нм (рис. 1.4).
Как уже было сказано выше, RyR-каналы располагаются на мембране СР; при переходе в открытое состояние через них происходит высвобождение ионов Са2+ из просвета ТЦ (люмена). Согласно данным электронной микроскопии [12], RyR-канал имеет форму четырехлистника со стороной 27 нм (рис. 1.4).

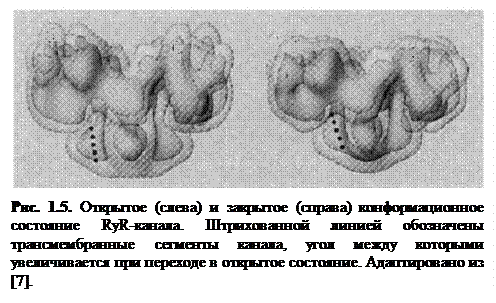
В процессе открытия RyR-канала изменяется его конформационное (структурное) состояние: данный белок изменяется по форме и структуре, переходя в проводящее состояние (рис. 1.5).
На рисунке 1.6 приведены результаты экспериментальных исследований структуры мембраны СР [13]; видно, что на мембранах RyR-каналы объединены в кластеры (решетки) (от 100 до 300 каналов в каждой). Каналы в решетке упорядочены и находятся под определенным углом друг к другу, также экспериментальные данные многих исследовательских групп говорят о существовании взаимодействия между каналами в кластере благодаря наличию между ними специфических связывающих белков [12-15].
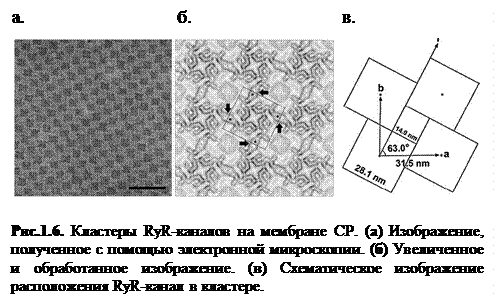
Из всего вышесказанного можно сделать следующие выводы:
· RyR-каналы имеют сложную молекулярную структуру и, соответственно, огромное количество степеней свободы.
· На мембране СР каналы образуют регулярную структуру решетки. Моделирование динамики RyR-каналов усложняется тем, что при разработке теории кластера RyR-каналов необходимо учесть взаимодействия между ними.
Date: 2015-08-24; view: 983; Нарушение авторских прав