Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Комплементарность
|
|
Например, чтобы человек имел нормальный слух, необходима согласованная деятельность нескольких пар генов, каждый из которых может быть представлен доминантным и рецессивным аллелями. У человека слух нормален, если каждый из них имеет хотя бы по одному доминантному аллелю в диплоидном наборе хромосом. В случае, если хотя бы один из них представлен гомозиготой рецессивной, человек будет глухим. Мы разберем это на простом примере, предположив., что нормальный слух формирует пара генов, и рассмотрим явление комплементарности. В брак вступает пара глухих, у них рождаются дети, которые все слышат. Это может быть только в случае, если один родитель страдает глухотой по гену «в» (ААвв), а другой — по гену «а» (ааВВ).
Р ♀ ААвв х ♂ааВВ
глухая глухой
Гаметы: Ав; аВ;
F1 АаВв;
слышащий
Все дети получат с гаметами Ав и аВ доминантные аллели, их генотипы будут одинаковы — АаВв, то есть каждая гамета будет иметь доминантный аллель по каждому гену, которые будут взаимодействовать, дополняя друг друга и формировать новый, по отношению к родителям, признак — нормальный слух. Это и есть комплементарное взаимодействие генов.
Допустим, что дети вступят в брак с себе подобными, и оценим вероятность рождения слышащих и глухих детей в этом случае.
Р ♀АаВв х ♂АаВв
слышащая слышащий
| ♂ ♀ | 1/4 АВ | 1/4 Ав | 1/4 аВ | 1/4 ав |
| 1/4 АВ | 1/16 А-В-слышащий | 1/16 А-В-слышащий | 1/16 А-В-слышащий | 1/16 А-В слышащий |
| 1/4 Ав | 1/16 А-В-слышащий | 1/16 А-вв-глухой | 1/16 А-В-слышащий | 1/16 А-вв глухой |
| 1/4 аВ | 1/16 А-В- слышащий | 1/16 А-В-слышащий | 1/16 ааВ-глухой | 1/16 ааВ-глухой |
| 1/4 ав | 1/16 А-В-слышащий | 1/16 А-вз глухой | 1/16 ааВ-глухой | 1/16 аавв глухой |
Таким образом, мы получили соотношение:
9 А-В-: 3 А-вв: 3 ааВ-: 1 аавв
слышащие глухие глухие глухой
или 9 слышащих: 7 глухих
Соотношение фенотипических классов при комплементарном взаимодействии может быть разным, в зависимости от вида комплементарности и проявляемости отдельных генов: 9:3:3:1; 9:6:1; 9:7.
8.2. Эпистаз
Эпистаз — подавление действия одного гена действием другого гена. Различают эпистаз доминантный (доминантный аллель эпистатирует, или подавляет проявление доминантного аллеля другого гена) и рецессивный (рецессивный аллель эпистатирует, ингибирует или подавляет проявление доминантного аллеля другого гена). Анализ при эпистатическом взаимодействии ведется так же на основе дигибридного скрещивания, как и в предыдущих случаях. Цифровые расщепления при эпистазе 12:3:1 (если каждый из двух генов имел самостоятельное проявление, при этом в первом поколении признак особей повторяет фенотип одного из родителей) и 13:3 (ген-ингибитор не имеет самостоятельного проявления, при этом фенотип родителей и первого поколения одинаков). Подавляющий ген называется геномсупрессором, или ингибитором, а подавляемый — гипостатическим. Этот тип взаимодействия наиболее характерен для генов, участвующих в регуляции работы генов в онтогенезе и работе иммунных систем человека (см. раздел 13.5).
Известно немало примеров эпистатического взаимодействия локусов у человека, приводящих к тому, что тот или иной доминантный аллель у некоторых индивидуумов не получает фенотипи-ческого выражения. Подобным примером может служить полидактилия, которая наследуется, как правило, по доминантному типу. Иногда встречается у детей «совершенно здоровых» родителей. В этом случае предполагается, что действие данного аллеля у родителей подавлялось другими генами (см. раздел 11.4).
8.3. Полимерия
Полимерия — обусловленность определенного (обычно количественного) признака несколькими эквивалентными (полимерными) генами. При таком взаимодействии индивидуальное проявление каждого усиливается в результате взаимодействия. Полимерия бывает двух видов'
некумулятивная — когда не важно количество доминаитных генов в генотипе, а важно его присутствие;
кумулятивная — когда число доминантных аллелей влияет на степень выраженности данного признака
У человека по типу кумулятивной полимерии наследуется пигментация кожи: чем больше доминантных аллелей, тем больше меланина образуется и тем интенсивнее окраска кожи. Разберем следующий пример, в котором необходимо определить вероятность появления детей с разными фенотипами в браке мулатов (особи дигетерозиготны).
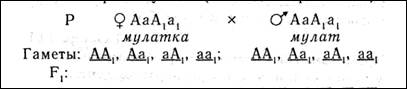
| ♂ ♀ | 1/4 АА, | 1/4 Аа, | 1/4 аА, | 1/4 аа, |
| 1/4 АА, | 1/16 АААА негр | 1/16 АААа светлокожий негр | 1/16 АААа светлокожий негр | 1/16 АаАа мулат |
| 1/4 Аа, | 1/16 АААа светлокожий негр | 1/16 ААаа мулат | 1/16 АаАа мулат | 1/16 Аааа светлокожий мулат |
| 1/4 аА | 1/16 АаАА светлокожий негр | 1/16 АаАа мулат | 1/16 ааАА мулат | 1/16 ааАа светлокожий мулат |
| 1/4 аа | 1/16 АаАа мулат | 1/16 Аааа светлокожий мулат | 1/16 ааАа светлокожий мулат | 1/16 аааа белый |
— негры (АААА) 1/16,
— светлокожие негры (АААа) 4/16,
— мулаты (ААаа) 6/16,
— светлокожие мулаты (Аааа) 4/16,
— белые (аааа) 1/16.
Обратите внимание, что при полимерии гены, как правило, обозначаются одинаковыми буквами, чтобы подчеркнуть однонаправленность их действия, т.к. при формировании признака не важно, какому гену принадлежат доминантные гены, а важно их количество.
Статистическое распределение вариантов цвета кожи оказывается непрерывным, т.е. аналогичным распределению роста и других метрических признаков (рис. 8.1).
В случаях наследования двух полимерных генов цифровые расщепления при скрещивании гибридов будут иметь четкие зависимости, являющиеся модификациями все того же расщепления для двух генов 9:3:3:1. При кумулятивной полимерии соотношение фенотипов будет 1:4:6:4:1, а'при некумулятивной полимерии — 15:1. В представленном выше примере, если допустить, что нас интересует факт наличия пигментации у человека, а не ее интенсивности, получим соотношение, характерное для некумулятивной полимерии: 15 пигментированных к 1 белому.
Если учесть, что спонтанные мутации происходят крайне редко (вероятность спонтанной мутации одного аллеля составляет 1/1000000-1/1000000000), то вероятность рождения у белых людей даже светлокожего мулата, а у негров — детей с более светлой кожей, крайне ничтожна.
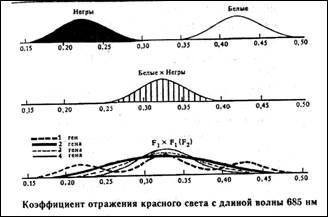
Рис. 8.1. Распределение цвета кожи у негров и людей белой расы. Цвет кожи оценивали по величине отражения света с длиной волны 685 нм. Кривые F2 представляют собой теоретически ожидаемые на основе различных предположений о числе генов, определяющих различие в цвете кожи между неграми и белыми. Исследование потомства F2 выявляет распределения, сходные с распределениями, полученными на основе предположения об участии трех или четырех пар генов [по W.F.Bodmer, L.L.Cavalli-Sforza (1976). Genetics, Evolution and Man, Freeman, San Francisco].
Меланин — пигмент, определяющий все мно-гобразие окрасок покровов, синтезируется в организме из аминокислоты тирозина в несколько этапов (рис. 8.2). Формирование промежуточного продукта диоксифениланина (ДОФА) — медиатора симпатической нервной системы свидетельствует о возможном плейотропном влиянии генов, участвующем в синтезе меланина, на свойства нервной системы. Меланин, формируя окраску покровов, является протектором от воздействия на организм УФ-лучей. Существуют два типа меланинов:
эумеланин — черный и темно-коричневый; феумеланин — желтый и рыжий.
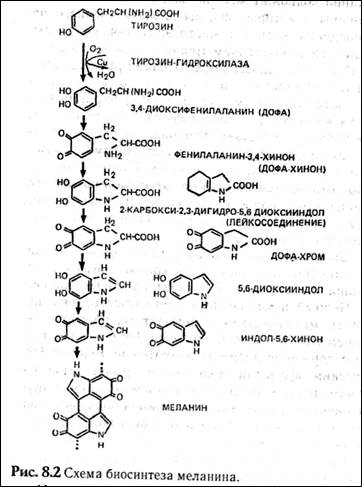
Показано, что на их синтез оказывает влияние метаболизм в клетках: при усиленном митозе клеток в основании волоса образуется феумеланин, а при замедлении эумеланин. При образовании фёумеланина важным условием является повышенная концентрация сульфгидрильных соединений, Феумеланин содержит меньше мономеров в молекуле. Описаны некоторые формы злокачественного перерождения клеток кожного эпителия, сопровождающиеся накоплением меланина (меланомы).
Все цвета волос, за исключением рыжих, составляют непрерывный ряд от темного до светлого, что соответствует уменьшению концентрации пигмента меланина, и наследуются полигенно. Есть основания утверждать, что различия в цвете волос обусловлены чисто количественными изменениями в содержании пигмента. Пигмент рыжих волос отличается по составу от меланина (феумеланин). Цвет волос часто меняется с возрастом, считают, что с наступлением половой зрелости пигментация стабилизируется.
Цвет радужной оболочки глаза зависит, с одной стороны, от присутствия гранул пигмента меланина, а с другой — от характера отражения света. Черный и коричневый цвета обусловлены наличием многочисленных пигментных клеток в переднем слое радужной оболочки. При светлых глазах содержание пигмента значительно меньше. Оптическим эффектом объясняется преобладание голубого цвета в свете, отраженном от депигменти-рованного переднего слоя радужной оболочки. Различное содержание пигмента определяет весь диапазон цвета глаз.
Следует помнить, что при кажущемся нарушении закона Менделя о независимом наследовании признаков для двух генов, связанного с тем или иным видом взаимодействия генов, в основе «неожиданных» цифровых соотношений лежит классическое расщепление 9:3:3:1. Кроме того, в каждом
конкретном случае необходимо понять, как происходит объединение расщепляющихся классов в единые фенотипы, только после этого можно интерпретировать результаты. Термин «взаимодействие генов» довольно условен, поскольку взаимодействуют не сами гены, а их продукты.
8.4. Гены-модификаторы
Генами-модификаторами называют такие гены, которые регулируют активность других генов, усиливая или ослабляя их проявления. В таком случае получается, что в формировании признаков, кроме «основных» генов, принимают участие гены-модификаторы. Такой тип наследования встречается очень часто. Примером модифицирующего аллеля у человека может служить брахидактилия. Существуют различные формы проявления: резкое укорочение указательного пальца и незначительное укорочение. Большинство больных с брахидак-тилией гетерозиготны (ВЬ). Анализируя родословную, можно предположить наличие гомозиготного рецессивного аллеля n-модификатора, определяющего крайнее проявление гена, и доминантного аллеля N — определяющего слабое проявление. Исходя из этого, генетическая конституция у индивида с выраженной брахидактилией будет Bbnn, у индивидов со слабой брахидактилией BbNn либо BbNN, а нормальными могут быть индивиды, принадлежащие к трем различным генотипам: bbnn; bbNn; bbNN (Штерн, 1965). Гены-модификаторы у людей выявляются очень сложно. Поэтому признание их наличия не помогает предсказать специфический характер проявления признака в будущих поколениях. И все же важно знать о тех
осложнениях, которые могут быть вызваны генами-модификаторами.
Природа генов-модификаторов до сих пор не выяснена: то ли это истинные гены-модификаторы, влияющие на действие основных генов, или это эффект плейотропии.
8.5. Плейотропия
Явления взаимодействия генов указывают на целостность генотипа при индивидуальном развитии особи, а признание принципа взаимодействия допускает, что все гены так или иначе взаимосвязаны в своем действии. Исходя из этого, мы можем сказать, что и один ген может влиять на несколько признаков. Такое множественное действие гена называют плейотропией. При плейотропии гены взаимодействуют на уровне продуктов контролируемых ими реакций, т.е. каждый ген ответствен за определенный этап метаболизма. Нарушения метаболизма на каком-либо из этапов отразятся на последующих (т.е. на нескольких элементарных признаках). У человека синдром Марфана (впервые описан в 1896 г.) обуславливается аутосомно-доминантным аллелем с высокой пенетрантностью и различной экспрессивностью. Синдром Марфана связан с высоким ростом, значительным удлинением костей пальцев ног и пальцев рук (паучьи пальцы — арахнодактилия), гиперподвижностью суставов, подвывихом хрусталика и аневризмом аорты. Соотношение полов равное, популяционная частота 0.04 на 1000 (Мак-Кьюсик). Такого рода аномалии скелета часто сопровождаются ненормальным расположением хрусталиков и пороками сердца. Разные симптомы, по Мак-Кьюсику,
Date: 2015-09-02; view: 1258; Нарушение авторских прав