Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Глава 4 хромосомы
|
|
4.1. Упаковка генетического материала
Если всю ДНК одной клетки вытянуть в одну линию, то ее длина была бы 1,74 м. Представленной в виде линии суммарной ДНК одного человека можно три раза обернуть земной шар по экватору. Поэтому хромосомы ядер клеток должны представлять собой сильно конденсированные структуры ДНК. При конденсации происходит уменьшение продольных размеров молекулы ДНК в десятки тысяч раз за счет образования сверхспиралей ДНК.
У эукариотических организмов значительная часть ДНК окружена множеством различных белков. Эти белки вместе с ДНК образуют комплексную структуру — хроматин, который обеспечивает специфический для эукариот тип регуляции экспрессии генов. В состав хроматина входят очень длинные двухцепочечные молекулы ДНК и белки гистоны, кислые белки и небольшое количество РНК- Всего известно пять типов гистонов: HI, H2A, Н2В, НЗ, Н4. Гистоны объединяют несколько групп основных белков. HI наиболее слабо свя-
зан с хроматином. Ядра нуклеосом состоят из гис-тонов 4-х классов: Н2А, Н2В, НЗ, Н4. Высокая консервативность свидетельствует об идентичности функций этих белков у всех эукариот.
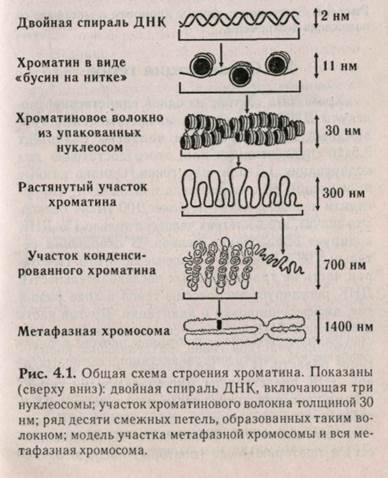
В хромосоме ДНК с помощью гистонов упакована в специальные регулярно повторяющиеся структуры — нуклеосомы. Так образуется структура, похожая на бусы, где каждая бусина — нук-леосома (диаметр около 10 нм). Нуклеосома представляет собой сегмент ДНК длиной около 200 пар оснований, навитый на белковую сердцевину, состоящую из восьми молекул белков-гистонов. Нук-леосомный кор (минимальная нуклеосома) «маскирует» 146 пар оснований ДНК. В нуклеосом-ную сердцевину (нуклеосомный кор) входит по две молекулы гистонов Н2А, Н2В, НЗ, Н4. Поверхности этих белковых молекул несут положительные заряды и образуют стабилизирующий остов, вокруг которого может закручиваться отрицательно заряженная молекула ДНК. Гистон HI размещается на участках ДНК, соединяющих одну нуклео-сому с другой. ДНК этих участков называют соединительной, или линкерной. Предполагают, что HI, возможно, регулирует транскрипционную активность хроматина и не участвует в стабилизации структуры хромосомы. Нуклеосомы, в свою очередь, упакованы в фибриллы, которые располагаются в хромосоме в виде хроматиновых петель. Хроматида построена из хроматиновых петель. Способ упаковки хроматина представлен на рис. 4.1.
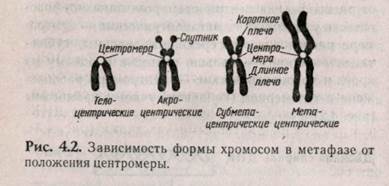
Митотическая хромосома состоит из двух сестринских хроматид и центромеры. В зависимости от расположения центромеры хромосомы морфологически разделяют на метацентрические — центромера расположена посередине хромосомы, субметацентрические — больше смещена к одному из краев и телоцентрические — центромера расположена в теломерном (концевом) участке хромосомы (рис. 4.2).
|
|
4.2. Организация генов
Хромосома состоит из одной единственной молекулы ДНК, содержащей множество генов. Гаплоидный набор хромосом человека содержит 3,5х109 нуклеотидных пар, этого достаточно для кодирования 1,5 млн. пар генов. Однако данные по изучению генома человека показывают, что организм человека имеет не более 100 тысяч белков. Это значит, что в клетках человека только 1% ДНК кодирует образование белков. В отношении остальных 99% существуют разные гипотезы. Некоторая часть не транслируемых последовательностей ДНК регулирует экспрессию генов в ходе развития, дифференцировки и адаптации. Другая часть избыточной ДНК входит в состав интронов, неко-дирующих участков генов, разделяющих кодирующие области. И все же большая часть избыточной ДНК представлена многочисленными семействами повторяющихся последовательностей. ДНК генома эукариот можно разделить на два класса последовательностей: уникальные (неповторяющиеся) и повторяющиеся (повторы) последовательности ДНК. К первому относятся однокопийные гены, кодирующие белки. Класс повторяющихся последовательностей ДНК представлен повторами, которые встречаются у человека от 2 до 107.
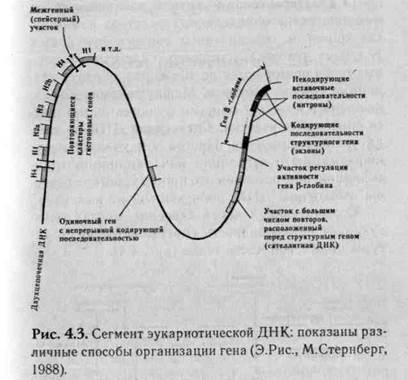
У человека выделяют структурные гены, функционирование которых тесно связано со специфическими последовательностями в молекуле ДНК, называемые регуляторными участками (рис. 4.3).
|
Структурные гены подразделяют на независимые гены, повторяющиеся гены, кластеры генов, прерывистые гены:
независимые гены — транскрибируются независимо, их транскрипция не связана с другими генами. Однако их активность может регулироваться, например, гормонами;
повторяющиеся гены — в хромосомах один ген может находиться в виде повторов, повторяясь много сотен раз, вплотную следуя друг за другом, образуя тандемы. Пример, гены рРНК;
кластеры генов — группы различных генов, находящиеся в определенных участках или локу-сах хромосом, объединенных общ ими функциями. В геноме человека, например, кластеры гистоно-вых генов повторяются до 10—20 раз, образуя тан-демные группы повторов. Между генами, объединенными в кластере общими функциями, находятся спейсерные участки. Спейсерная ДНК не всегда транскрибируется. Иногда эти участки несут информацию о регуляции или инициации транскрипции, но в основном это просто короткие повторы избыточной ДНК, роль которой не выяснена.
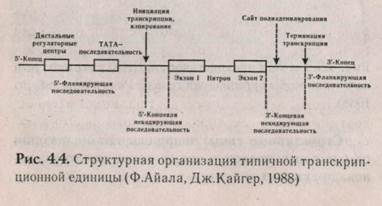
Отличительная черта строения многих генов эукариот — мозаичность (прерывистость) структуры смысловой части генов (рис. 4.4).
|
Это отличие связано с чередованием смысловых участков, несущих информацию о последовательности аминокислот в белке — экзонов, и участков некодирующих последовательностей, которые по длине значительно больше экзонов — интронов. В результате общая длина гена оказывается больше, чем можно было ожидать, примерно, в 5-7 раз.
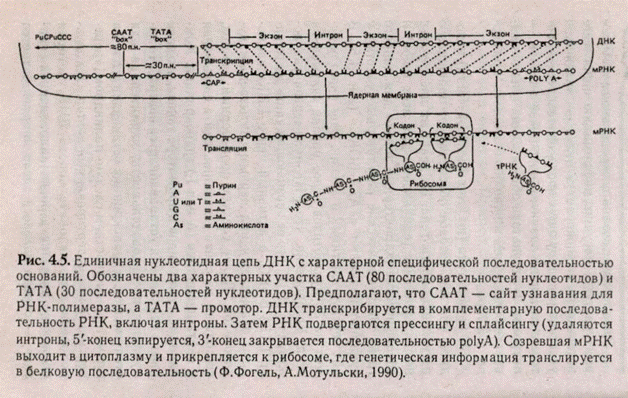
В начале гена (до его смысловой части) находятся участки, которые обеспечивают правильную регуляцию работы гена. Их делят на две группы: неспецифические (одинаковые для всех генов) и специфические (характерные только для данного гена). Неспецифические регуляторные участки у эукариот называют «TATA-БОКС» (рис. 4.5). В них чередуются тимин и аденин. Этот участок лежит на 30 нуклеотидов левее начала считывания гена. Установлено, что РНК-полимераза так ложится на ДНК, что ее опознающая часть закрывает TATA-БОКС, а активный центр РНК-полиме-разы оказывается над первым считываемым ну-клеотидом. Далее следует промоторный участок, состоящий из сайта-узнавания, сайта-связывания и сайта-инициации (А или Г) транскрипции. Комбинация нуклеотидов в промоторе такова, что если рамка считывания устанавливается неверно, то получается последовательность, характерная для стоп-кодонов, и транскрипция останавливается. За промоторным участком следует палиндром («перевертыш»), или инвертированный повтор. Это двух-цепочечная последовательность ДНК, которая одинаково читается в обоих направлениях («казак» — справа налево и слева направо читается одинаково). Иногда такую последовательность называют «областью с двойной симметрией», где осью симметрии является центральная точка, относительно которой последовательность остается одинаковой. Важное свойство палиндромов — возможность образовывать шпильки в РНК или структуры креста — в ДНК.
Если рамка считывания установлена неверно, то палиндром ДНК превращается в «крест», что делает невозможным дальнейшее продвижение фермента. Иногда палиндромы могут находиться в конце гена после его смысловой части и также, образуя «крест», будут прекращать синтез мРНК. Смысловая часть гена, как мы уже писали выше, состоит из чередующихся интронно-эк-зонных последовательностей. В структурной части гена расположены промотор и палиндром.
В последнее время выделяют включения специфической регуляции гена — энхансеры. Они расположены впереди гена на расстоянии "в сотни и тысячи нуклеотидных пар. У эукариот существуют специальные регуляторные белки, опознающие энхансер и присоединяющиеся к нему. Таким образом происходит активизация гена.
Геном образован не только ядерными, но и цитоплазматическими генами. В геноме гены выполняют разные функции. В зависимости от выполняемых функций гены подразделяют на обязательные гены, они кодируют жизненно важные функции в клетках всех типов (иногда их называют генами «домашнего хозяйства»), структурные гены, регуляторные гены,
пунктуационные гены.
4.3. Дополнение. Геномная дактилоскопия
В 1987 г. впервые была проведена идентификация личности по анализу ДНК (геномная дактилоскопия). Это стало возможным благодаря открытию Элика Джеффриза в 1985 г., связанному с обнаружением у генов человека структурного полиморфизма определенных тандемов, которые образуют «картину», специфичную для молекулы ДНК конкретного человека.
Тандемные последовательности состоят из многократно повторенных единиц, чаще богатых ГЦ парами. Частота повторов варьирует у различных индивидуумов от 14 до 300 раз.
Процесс идентификации ДНК сложен, но высокоэффективен. Процесс начинается с выделения ДНК, с последующей ферментативной обработкой специфическими рестриктазами, которые разрезают ДНК на отрезки. Отрезки группируют по размерам, а потом их идентифицируют радиоактивными маркерами — зондами. Зонды искусственно синтезируют и радиоактивно метят по фосфору, они содержат определенные последовательности (ми-нисателлиты), богатые ГЦ нуклеотидными парами. Таким образом, используя один зонд, можно наблюдать за наследованием большого количества аллелей (рис. 4.6).
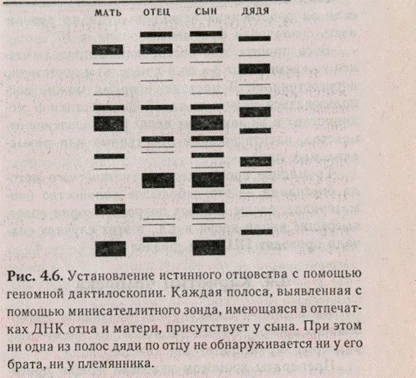
Получаемый сложный набор вариабельных полос и заключает в себе специфическую картину геномного «дактоотпечатка» (DNA fingerprint), аллели, формирующие эту картину, стабильны в соматических и зародышевых клетках и наследуются в соответствии с законами Менделя.
Это метод часто применяется для установления отцовства, т.к. практически все полосы геномного отпечатка данного человека должны выявляться в отпечатках либо его матери, либо отца. Около половины всех полос должны иметь отцовское происхождение. Случайное появление или отсутствие полос маловероятно. Если предположить, что скорость мутирования равна Ю-4 т.п.н., то при использовании всех трех проб разрешению поддаются в среднем 15 фрагментов отцовского происхождения в диапазоне длин 4-20 т.п.н. Вероятность того, что предполагаемый отец будет случайно иметь 15 фрагментов < 4 х 10~п, в случае,
если он родственник истинного отца, то вероятность совпадения фрагментов равна < 10~5.
Весь процесс диагностирования продолжителен во времени (от 3-х до 4-х недель) и достаточно дорогостоящий. В настоящее время можно воспользоваться услугами такой диагностики в медицинских и юридических целях (установление отцовства, идентификация преступных или разыскиваемых лиц).
Громадное преимущество генетического метода опознания — это небольшое количество биоматериала: следы кожных покровов, корни волос, высохшие капли крови и т.д., в этих случаях сначала проводят ПЦР (см. раздел 3.4).
4.4. Кариотип человека
Кариотип — совокупность признаков хромосомного набора: форма хромосом, их количество, размеры, характерные для каждого вида.
Препараты хромосом человека можно приготовить из всех тканей и клеточных суспензий, содержащих делящиеся клетки. Но чаще препараты метафазных хромосом готовят из лимфоцитов периферической крови, которые предварительно культивируют в присутствии митогена (вещество, способное индуцировать митоз, в частности, фи-тогемагглютинин). Этот метод был предложен Мурхедом с соавт. в 1960 г.
Классификация и номенклатура равномерно окрашенных хромосом человека была выработана на международных совещаниях, созывавшихся в Денвере (I960), Лондоне (1963) и Чикаго (1966). Согласно рекомендациям этих конференций, хро-
мосомы располагаются в порядке уменьшения их длины. Все хромосомы разделены на семь групп, которые были обозначены буквами английского алфавита от А до G. Все пары хромосом было предложено нумеровать арабскими цифрами.
Группа А (1—3) — самые крупные хромосомы. Хромосомы 1 и 3 — метацентрические, 2 — суб-метацентрическая. Группа В (4—5) — две пары довольно длинных субметацентрических хромосом. Группа С (6—X—12) — хромосомы средних размеров. Хромосомы 6, 7, 8 и 11 больше похожи на метацентрики (центромерный индекс 40-30%). Хромосомы 9, 10 и 12 — субметацентрики. Х-хромосома по размеру и морфологии сходна с хромосомами 6 и 7. Группа D (13—15) — акроцентрические хромосомы средних размеров. Группа Е (16— 18) — довольно короткие хромосомы. Хромосома 16 более метацентрична, часто на промаксималь-ном конце длинного плеча имеется вторичная перетяжка. Группа F (19—20) — самые маленькие метацентрики, практически между собой не различимы. Группа G (21—22) — две пары самых мелких акроцентрических хромосом. Y-хромосома выделяется как самостоятельная (рис. 4.7, 4.8).
При этом хромосомы различных групп хорошо отличаются друг от друга, в то время как внутри группы их невозможно различить, за исключением группы А.
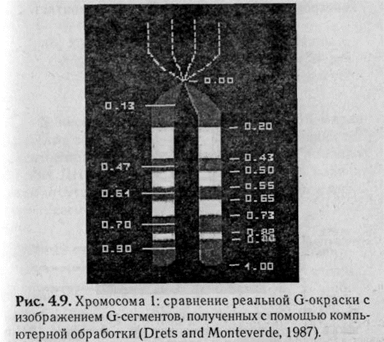
В начале семидесятых годов был разработан метод дифференциальной окраски хромосом, который позволил идентифицировать каждую из хромосом. На рис. 4.9 показана хромосома 1-го человека с изображением дифференциального окрашивания и нумерации сегментов.

Каждая хромосома человека содержит только ей свойственную последовательность полос, что позволяет точно идентифицировать каждую хромосому и с более высокой точностью определить, в каком сегменте произошла перестройка. Поперечная исчерченность хромосом есть результат неравномерной конденсации гетеро- (высокоспирализованная ДНК) и эухроматина (релаксиро-ванная ДНК) на протяжении всей длины хромосомы, отражающий порядок расположения генов в молекуле ДНК
Хромосомы в метафазе максимально конденсированы, в профазе и прометафазе более расплетенные, что позволяет выявлять еще большее количество сегментов — 800-1200, некоторые авторы описывают более 2000. Этот метод используют для более точной идентификации точек разрывов хромосом и микроделеций (рис. 4.10).
На метафазной хромосоме.(рис. 4.10, 1М) слева приведены символы, которыми принято обозначать короткие (р) и длинные (q) плечи хромо- I сом, а также номера расположения сегментов. Эти обозначения используются при описании карио-типа. Сначала указывают общее число хромосом и набор половых хромосом, затем отмечается, какая хромосома лишняя или какой не хватает.

Указывается номер хромосомы и обозначается плечо, в котором произошли изменения (символы хромосомных аберраций см. в гл. 12), после написания символа плеча указывается номер сегмента. Например:

В настоящее время существуют ДНК-маркеры (или зонды) для многих еще более мелких сегментов практически всех пар хромосом. С помощью таких ДНК-зондов можно точно оценить наличие или отсутствие определенного, даже очень маленького, сегмента в хромосоме (рис. 4.11).

4.5. Дополнение. Феномен ассоциаций акроцентрических хромосом человека
Среди разновидностей неслучайного расположения хромосом в метафазе выделяют феномен центромерных ассоциаций (рис. 4.12).
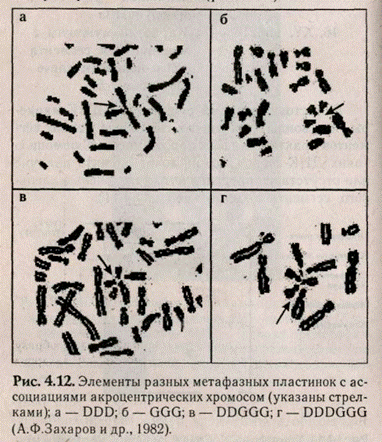
Феномен ассоциации акроцентрических хромосом (ААХ) был впервые описан в 1961 г. M.A.Fer-gusson-Smith and S.D.Handmaker и получил название «спутничная ассоциация». Способность акроцентрических хромосом вступать в спутничные ассоциации была подтверждена многими исследователями (Прокофьева-Бельговская А.А. и др., 1969; Захаров А.Ф. и др., 1982 и др.).
Феномен ААХ привлекает пристальное внимание в связи с ядрышкообразующей функцией акроцентрических хромосом. С активностью ядрышка связывают процесс синтеза рРНК, из которой впоследствии формируются в цитоплазме рибосомы (рис. 4.13), а также и другие важные процессы функционирования ядра,.такие как транспорт различных видов РНК из ядра в цитоплазму, поэто+ му ААХ рассматривают как специфический показатель структуры и функции интерфазного ядра.

Уже в первых исследованиях было установлено, что феномен ААХ связан с ядрышкообразующей функцией акроцентрических хромосом в профазе митоза, когда еще сохраняется ядрышко и спирализация хромосом только начинается, остаточное ядрышко обычно связано со спутничными
районами коротких плечей акроцентрических хромосом. Позже было показано, что короткие плечи всех 10 акроцентрических хромосом человека содержат 3 сегмента: Р1 — проксимальная часть, прилегающая к центромере; Р2 — спутничная нить, в районе которой располагается ядрышковый организатор (ЯОР); РЗ — маленькое гетерохроматическое тельце, содержащее теломеру короткого плеча (Paris Conferens, 1971).
Одна из первых гипотез, объясняющих природу ААХ, была сформулирована А.А.Прокофьевой-Бельговской (1966): специфическая ориентация акроцентрических хромосом является следствием соматической конъюгации гомологических участков. Конъюгация происходит за счет относительной гомологии амплифицированных участков ДНК, расположенных в коротких плечах ААХ. Благодаря этому свойству гетерохроматиновых районов в интерфазном ядре формируются хромоцентры из гомологичных и негомологичных хромосом, что способствует определенной их упорядоченности в интерфазном ядре, которые сохраняются во время митоза в виде неслучайного расположения в ме-тафазной пластинке.
Если гипотеза А.А.Прокофьевой-Бельговской предполагает, что «формирование общего ядрышка представляет собой не причину ассоциаций, а следствие этого процесса», то противоположную точку зрения имеют авторы «нуклеолярной гипотезы» M.A.Fergusson-Smith and S.D.Handmaker. Они полагают, что если ядрышки в интерфазном ядре имеют тенденцию к слиянию, то этот процесс приводит к ААХ. По мнению многих исследовате-
лей, частота ядрышек в интерфазе высоко коррелирует с числом ААХ (Гиндилис В.Я., 1967; Ми-кельсаар А.В., 1979 и др.).
Данные последних лет о химическом составе хромосом, их молекулярной организации, строе нии и функции ЯОР акроцентрических хромосом человека позволяют сделать заключение, что «конъ-югационная» и «нуклеолярная» гипотезы о природе ААХ не взаимоисключают, а взаимодополняют друг друга.
По мнению исследователей, частота ААХ в популяции лимфоцитов является более информативным показателем, чем аберрации хромосом, так как она отражает все процессы жизнедеятельности клеток. Образование ААХ в интерфазе является нормальным и физиологическим процессом. Частота ААХ в лимфоцитах тесно связана с их иммунореактивностью, что постоянно наблюдается при развитии иммунных реакций, так как этот показатель характеризует пролиферативную активность лимфоцитов (Фролов А.К. и др., 1993). Все это позволяет широко применять метод учета ААХ как иммуноцитогенетический тест.
4.6. Задание
1. Что такое хромосома? хроматида? хромонема? хроматин? хромомера?
2. На каких стадиях митоза хромосомы хорошо видны? Ответ обоснуйте.
3. Что такое кариотип? Выберите правильный ответ: а) совокупность всех признаков и свойств организма; б) совокупность хромосом, характерных для данного вида; в) совокупность всех генов организма.
4. По какому признаку отличаются кариоти-пы мужчины и женщины: а) по количеству ауто-сом; б) по количеству гетерохромосом; в) по форме гетерохромосом; г) по гетерохромосомам; д) по соотношению аутосом и Х-хромосом?
5. Могут ли быть в кариотипе человека 30 хромосом отцовскими? Ответ поясните.
6. Объясните, чем различаются между собой следующие кариотипы: 47, XY, +21; 46, ХХ,-р21; 46, XX; 46, XY,+q21.
Date: 2015-09-02; view: 2478; Нарушение авторских прав