Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Загальна характеристика черепа рибоподібних і риб 1 page
|
|
Череп — це складний конгломерат кісток різної будови, функції і походження, об'єднаних у єдине морфо-функціональне ціле.
Складність будови черепа визначається різноманіттям властивих йому функцій, а значна розмаїтість його будови — специфічними відмінностями частини цих функцій у різних риб. Череп утворює ряд захисних капсул для життєво важливих органів: головного мозку і органів нюху, зору, слуху і рівноваги. Частиною черепа є щелепний апарат, а також ряд органів функціонально зв'язаних з цим апаратом.
Характерною рисою черепа, що відрізняє його від інших частин скелета, є приналежність більшості його кісток до первинних або покривних кісток.
Анатомічно череп поділяють на три природно відокремлені відділи — мозковий череп, нижню щелепу і під'язикову кістку. У мозковому черепі, у свою чергу, розрізняють мозковий і лицьовий відділи. Перший містить у собі головний мозок, орган рівноваги і слуху і частково органи нюху і зору. Другий — бере участь в утворенні ротової порожнини з щелепним апаратом.
У черепі хребетних, орієнтуючись на його розвиток у філогенезі та онтогенезі, розрізняють два відділи — нейрокраній або осьовий череп і спланхнокраній або вісцеральний череп. Обидва ці відділи в хребетних виникають і перший час розвиваються як утворення зовсім різного пристосувального значення. Нейрокраній з'являється в зв'язку з прогресивним розвитком переднього кінця нервової трубки, що перетворився у головний мозок, а вісцеральний череп — у передньому відділі кишкової трубки в результаті формування органів захоплювання їжі.
Серед сучасних хребетних найпримітивніший осьовий череп мають круглороті. їх череп у значній мірі сполучнотканинний перетинчастий.
Хрящ розташовується лише знизу, частково з боків і зверху. у хрящовій пластинці, яка лежить біля основи черепа, знаходиться значних розмірів основна фонтанель. У склепінні черепа хрящ представлений тільки вузьким містком, що лежить на рівні слухових капсул. Останні утворюють задній кінець черепа, але не включені в мозкову коробку, а лише приростають до неї з боків. Потиличної частини черепа в круглоротих ще немає. Спереду до черепа волокнистою тканиною приєднується непарна нюхова капсула (рис. 132).
Череп хрящових риб представляє собою хрящову коробку, що цілком охоплює головний мозок (рис. 133). Тільки в передньому відділі склепіння залишається досить велика верхня фонтанель, затягнута сполучнотканинною перетинкою. Нюхові і слухові капсули асимілюються стінками черепа, утворюючи, однак, попереду і позаду характерні здуття. За слуховими капсулами лежить потилична частина черепа, а перед нюховою ділянкою череп доповнюється хрящовим виступом, що лежить в основі рила {rostrum).
В ділянці очних ямок череп трохи стиснутий з боків, але відноситься до черепів платібазального типу, тобто до черепів із широкою основою, у яких між очними ямками знаходиться головний мозок. У тропібазальних черепах, або черепах з вузькою основою, що властиве рибам та деяким іншим хребетним, медіальні стінки очних ямок можуть бути зближеними до взаємного торкання, перетворюючись в міжочну перегородку {septum interorbitale) (рис. 134).

Рис. 133. Череп акули: 1 — губні хрящі; 2 — піднебінно-квадратний хрящ; З — гіомандибулярний хрящ; 4 — 1, 2, 3-я зяброві дуги; 5 — фарінгобран-хіале; 6 — епібранхіале; 7 — кератобранхіале; 8 — базібранхіале; 9 — гіпоб-ранхіале; 10 — гіоїд; 11 — базігіале; 12 — меккелів хрящ
Матеріалом для розвитку осьового черепа у хребетних служать склеротоми головних сомітів (рис. 135, 136). Розрізняють 7-9 пар головних сомітів. Три передніх (2-й та 3-й подвійні) лежать спереду від закладки вуха і називаються передвушними сомітами, інші — завушними (рис. 135).
Для головних сомітів характерний сильний розвиток склеротомів і слабкий міотомів. Це пояснюється необхідністю більшої кількості
скелетогенного матеріалу для розвитку черепа, ніж тулубового скелету, і втратою функції соматичною мускулатурою в ділянці черепа внаслідок зрощення окремих сегментів.
Мезенхіма, яка виникає із склеротомів головних сомітів, є джерелом сполучної тканини, що покриває суцільною оболонкою головний мозок разом із закладками органів чуття і хордою. Це стадія перетинчастого черепа, яка властива всім хребетним на ранніх стадіях розвитку.

Рис1. 135. Сегментація мезодерми в ділянці голови (за Алленом В. Ф., 1914 p.): 1 — головний мозок; 2 — нервова трубка; 3 — соміт; 4 — хорда; 5 — кишка; б — целом; 7 — епідерміс; 8 — глотка; 9 — зяброві щілини; 10 — очна капсула

Рис. 136. Поперечний розріз через голову зародка хребетного: 1 — мозок; 2 — гангліонарна пластинка; 3 — закладка осьового черепа; 4 — закладки вісцерального скелета; 5 — глотка; 6 — хорда; 7 — склеротом
Перетворення перетинчастого черепа в хрящовий відбувається подібно у всіх хребетних. У сполучній тканині, яка огортає мозок, з'являються місцеві згущення, що мають сталість розташування, у яких потім розвивається хрящова тканина.
Є декілька загальних для всіх хребетних закладок хрящового черепа. В основі черепа з'являється дві пари хрящів: з боків переднього кінця хорди розвиваються парахордальні хрящі (cartilagо parachordalia cranii), а спереду від них під переднім мозком — черепні балки (trabeculae cranii). У бічних стінках черепа закладаються очноямкові або клино-латеральні хрящі (cartilagines orbitaless. sphenolaterales). Навколо органів чуттів розвиваються хрящові капсули: нюхова, слухова та органу зору (рис. 137-138).
Парахордалії є продуктом зрощення первинно відособлених сегментів осьового скелета. Про це свідчить їхнє положення щодо хорди і наявність у хрящових риб слідів сегментації в їхньому задньому відділі у вигляді подвійної закладки в кожному сегменті верхніх дуг хребців (рис. 137). У відповідності з розвитком із цього відділу парахордалій потиличної частини черепа, його називають потиличним.
Передній відділ парахордалій, що лежить між вушними капсулами і не зберігає слідів сегментації, називають слуховим відділом (рис. 138). Усю частину неврального черепа, яка розвивається з парахо-далій, називають хордальним черепом і розглядають як видозмінений осьовий скелет.

Рис. 137. Розвиток хрящового черепа в зародка акули: 1 — трабекули; 2 — мозок; 3 — око; 4 — очноямковий хрящ; 5 — слухова капсула; 6 — парахордалії; 7 — хорда; 8 — зяброві дуги; 9 — під'язикова дуга; 10 — щелепна дуга
Його передня межа чітко позначена розташуванням гіпофіза.
Відділ неврального черепа, що походить із трабекул і очноямкових хрящів, називають прехор-дальним. Його, на противагу хор-дальному черепу, варто розглядати як нове утворення, що виникає в зв'язку з розвитком переднього мозку і відповідних органів чуття.
Подальший розвиток хрящового черепа відбувається шляхом збільшення розмірів і зрощення названих раніше первинних закладок. Звернені один до одного кінці парахордалій і трабекул зростаються, але так, що між ними ще довгий час залишається отвір, який називається основною фонтанеллю (fenestra basicranialis anterior). 3 переднім кінцем трабекул із самого початку тісно зв'язані нюхові капсули, а з передньою ділянкою парахордалій уже незабаром зростаються слухові капсули. Очна капсула завжди залишається відособленою, рухомою і розвивається в склеру ока.
Виникаюча в такий спосіб хрящова основа мозкового черепа починає розростатися в різні боки. Вона зливається при цьому з очноямковими хрящами і далі розростається з боків головного мозку. Передній кінець хрящової пластинки основи черепа проникає між нюховими капсулами, формуючи носову перегородку. Надалі хрящ розростається і на склепіння черепа, хоча майже ніколи його повністю не покриває. На ній залишаються затягнуті сполучною тканиною вікна.
Повністю хрящовим залишається дефінітивний мозковий череп і в хрящових ганоїдів. Він являє собою хрящову коробку, що охоплює головний мозок і охоплює нюхові і слухові капсули. Добре видно, що видовжений потиличний відділ черепа утворений шляхом приростання декількох хребців (рис. 139).
Сильно, хоча і неоднаково у різних представників цього ряду розвинуте рило. У старих риб у хрящі з'являються точки окостеніння.
Зверху і з боків хрящовий нейрокраній осетрових укритий суцільним панциром покривних кісток. У ньому є отвори тільки для ніздрів, очей і бризкалець. Розташування і форма цих кісток в осетрових ще не стали, хоча серед них є уже кістки, що нагадують покривні кістки вищих риб (рис. 140). Покривні кістки (леміш і па-расфеноїд) підстилають хрящовий череп осетрових також знизу (рис. 139).
У кісткових ганоїдів та кісткових риб хрящ в осьовому черепі частково витісняється кістками, що розвиваються на його основі. Є кістки змішаного походження. Більше ніж в інших місцях, хрящ зберігається в ділянці нюхової капсули, де його знаходять у кісткових ганоїдів.
Стабільними для риб із заміщуваних кісток є такі, як основна потилична (basioccipitale), бічні потиличні (os occipitalia lateraliа) і верхня потилична (os supraoccipitale), відсутня ще в кісткових ганоїдів; у основі черепа знаходяться ще основна клиноподібна (os basis-phenoideum) та передня клиноподібна (os praesphenoideum).

Рис. 139. Череп осетра збоку після видалення покривних кісток: 1 — гіоїд; 2 — інтергіалє; 3 — нижня щелепа; 4 — піднебінно-квадратний хрящ; 5 — симплектікум; 6 — гіомандибула; 7 — зяброві дуги;

Рис. 140. Дермальне склепіння черепа осетра: 1 — лобові кістки; 2 — тім'яні кістки
Остання відсутня у костистих риб (рис. 141). З боків від цих кісток в очноямковій ділянці лежать бічні клиноподібні кістки (ossa laterosphe-noidea) і очно-клиноподібні (ossa orbitosphenoidea). Останні є тільки в нижчих риб, у вищих риб вони зливаються в непарну міжочну перегородку. За рахунок окостеніння слухової капсули виникають передня вушна (os prooticum), задня вушна (os opisthoticum) і верхня вушна (os epioticum) кістки. Внаслідок вростання в хрящ покривних задньої лобової і надскроневої кісток у костистих риб розвиваються в цій же ділянці ще клиновушна (os sphenoticum) і за нею крило-вушна (os pteroticum), а в нюховій ділянці подібним чином виникають змішані кістки: непарна середня решітчаста (os mesethmoideum) і парна бічна решітчаста (os ectoethmoideum).
Покривні кістки у кісткових ганоїдів і костистих риб набувають настільки постійну форму і розташування, що дозволяють провести їхню гомологізацію у представників різних рядів. Вони витісняють осьовий череп тільки в ділянці його склепіння. З боків між зовнішнім панциром, утвореним цими кістками, та осьовим черепом залишаються великі простори, заповнені елементами вісцерального скелета і мускулатурою (рис. 142).
У кісткових ганоїдів розрізняють наступні покривні кістки: вздовж серединної лінії розташовуються тім'яні (ossa parietalia), лобові (ossa frontalia) і носові (ossa nasalia). До них спереду примикають верхня нюхова (os supraethmoideum) та передлобова (os prefrontak), а ззаду — задні тім'яні (ossa postparietalia). Очна орбіта оточена рядом орбітальних кісточок (ossa orditalia), а позаду них розташовані задня лобова (os postfrontale), надскронева (os supratemporale) і луската (os squamosum). Нижче очної орбіти і збоку на черепі знаходиться кілька щічних кісток (ossa jugalia) (рис. 143).

Рис. 141. Схема черепа костистої риби: I-V зяброві дуги; 1 — під'язикова кістка (os hioideum); 2 — суглобова кістка (os articulare); 3 — зубна кістка (os dentate); 4 — симплектикум {symplecticum); 5 — квадратна кістка (os quadratum); 6 — задня крилоподібна кістка (os postpterygoideum); 7 — зовнішня крилоподібна кістка (os ectopterygoideum); 8 — внутрішня крилоподібна кістка (os endopterygoideum); 9 — піднебінна кістка (os palatinum); 10 — верхньощелепна кістка (maxilla); 11 — передщелепна кістка (os ргаетахіНа); 12 — середня решітчаста кістка (os mesethmoideum); 13 — носова кістка (os nasale); 14 — бічна нюхова кістка (os ectoehtmoideum); 15 — очноямково-клиноподібна кістка (os orbitosphenoideum); 16 — бічна клиноподібна кістка (os laterosphenoideum); 17 — лобова кістка (os frontale); 18 — основна клиноподібна кістка (os basisphenoideum); 19 — клино-вушна кістка (os spher noticum); 20 — передня вушна кістка (os preoticum); 21 — крило-вушна кістка (os pteroticum); 22 — задня вушна кістка (os postoticum); 23 — тім'яна кістка (os parietale); 24 — верхня вушна кістка (os epioticum); 25 — верхня поти| лична кістка (os supraoccipitale); 26 — бічна потилична кістка (os occipitale laterale); 27 — основна потилична кістка (os basioccipitale); 28 — паракли* ноподібна кістка (os parasphenoideum); 29 — нижньощелепна кістка (man* dibula); ЗО — міжпід'язикова кістка (os interhiale)
У костистих риб відсутні задні тім'яні, лускаті і щічні кістки.
Інакше побудований нейрокраній у кистеперих риб, причому дуже подібно у целокантових представлених сучасною латимерією і у викопних рипідістій, що вважаються предками наземних хребетних. Нейрокраній у кистеперих риб складається з двох кісткових блоків, рухливо з'єднаних між собою, переднього — етмосфеноїдного (ethmosphenoideum) і заднього отикоокципітального (oticooccipitale) (рис. 144). Етмосфеноїдний блок включає нюхову і клиноподібну частини черепа та вміщає в собі орган нюху, органи зору і передній відділ головного мозку. Отикоокципітальний блок складається з потиличної кістки і слухових капсул та охоплює орган рівноваги і слуху і задній відділ головного мозку. Вважають, що етмосфеноїдний блок відповідає трабекулярному, а отикоокципітальний — пара-хордальному відділам ембріонального черепа гнатостомат.
Окремі окостеніння в обох блоках непомітні. Хрящ у різних рипідістій більш-менш зберігається тільки в ділянці нюхових капсул, у передній ділянці вентральної стінки основи отикоокципитального блоку й у основі вушних капсул.
Дотичні поверхні блоків формують зчленування. їх зв'язує, крім того, хорда, що пронизує основу отикоокципіталь-ного блоку та упирається в заглиблення на задньому кінці етмосфеноїда (рис. 145).
Покривні кістки в кистеперих риб відповідно блокам ней-рокранія, на яких вони розташовуються поділяють на два відділи або щити — етмосфе-ноїдний і отикоокципітальний (рис. 145-146).
Етмосфеноїдний щит рипідістій характеризується порівняно невеликим предочноям-ковим відділом, що складається з безлічі дрібних кісток, кількість яких зменшується від нижчих форм рипідістій до вищих; (рис. 145-146).

Рис. 144. Черепна коробка девонського целаканта (Nesides): 1 — отикоокципітальний блок; 2 — етмосфеноїдний блок; 3 — нюхова ямка; 4 — базип-терігоїдний відросток

Рис. 145. Черепна коробка сучасної латимерії: 1 — екзетмоїд; 2 — етмосфе^ ноїдний блок; 3 — отикоокципітальний блок; 4 — суглобові ямки для під'язиково-нижньощелепної кістки; 5 — отикоокципітальний відросток для з'єднання з етмосфеноїдом; 6 — хорда; 7 — базісфеноїд; 8 — парасфеноїд

Рис. 146. Кістки дермального склепіння черепа в примітивних остео-лепідин: 1 — тім'яна; 2 — пінеальний отвір; 3 — задні ростральні; 4 — середні задні ростральні; 5 — передня задня ростральна; 6 — носова, 7 — носорило-передверхньощелепна; 8 — тектальні передні; 9 — ніздря, 10 — латеральна ростральна; 11 — сьома носова; 12 — супраорбіто-тектальна; 13 — задня супраорбітальна; 14 — міжскронева; 15 — надскронева; 16 — задньотім'яні передні, 17 — задньот тім'яні задні; 18 — табличні
Кістки ці можуть досить різноманітно зливатися між собою. Передній край щита утворений парними передщелепними кістками (praemaxillarid), що зливаються з середньою ростральною (rostrate mediate) і передньою носовою (nasale anterior) у комплексну naso-rostro-praemaxillare. З заду до них прилягає латеральна ростральна кістка (os rostrate laterale), що обмежує латерально ніздрю. Медіально тягнеться серія кісточок, що позначаються як задні ростральні (ossa postrostralia). У ранніх остео-лепід кісточки цієї серії займають велику передню половину розглянутого щита, досягаючи ззаду до пінеального отвору (рис. 146). У їх числі розрізняють передню, середню і декілька задніх кісточок (os postrostrate anterior, medium et posterior). Останні у вищих форм представлені однією кісткою (рис. 147).
З етмосфеноїдним щитом латерально зв'язані верхньощелепна кістка і кістки щічної пластини, до яких відносяться слізна, вилична, заорбітальна, квадратновилична, луската і передкришкова.
Знизу етмосфеноїд покриває піднебінна пластинка передверхньо-щелепної кістки, леміш (vomer) і парасфеноїд.

Рис. 147. Дермальні кістки черепа еустеноптерона: 1 — надскронева; 2 — луската; 3 — міжскронева; 4 — задня кістка очної орбіти; 5 — шкірна клиноподібна; 6 — задня надорбітальна; 7 — сьома носова; 8 — постростральна задня; 9 — постростральна середня; 10 — постростральна передня, 11 — друга носова; 12 — тім'яна; 13 — задня тім'яна
Отикоокципітальний щит утворений трьома парами кісток. З боків серединної лінії лежать задні тім'яні кістки (ossa postparietalid), що у деяких ранніх форм можуть бути представлені передніми і заднімц. Латерально від них знаходяться надскронева (os supratemporale) і таблична (os tabulare). Передній край щита співпадає з цим же краєм відповідного блоку нейрокранія, а його задній край знаходиться на; рівні межі вушної і потиличної ділянок.
Вісцеральний скелет з'явився раніше і незалежно нейрокранія.1 У сучасного ланцетника він вже представлений понад 100 стрижня-' ми з безклітинної опорної речовини, що лежать у зябрових перегородках, з кожного боку.
Розвиток щелепноротих був пов'язаний з надбанням активного!, розширення зябрової порожнини, яке досягалося дією на вентральні і дорсальні кінці вісцеральних дуг гіпаксональною соматичною мускулатурою, що одержала прикріплення на них, тобто шляхом розвитку епібранхіальної і гіпобранхіальної мускулатури.
Активне розширення зябрової кишки, що виникало таким шляхом в силу дії соматичної мускулатури і стискання вісцеральною мускулатурою, значно збільшили розмах зміщення її стінок і спричинили поділ зябрових дуг на 4 членики: глоткозябровий — pharyn-gobranchiale, верхній зябровий — epibranchiale, ріжковозябровий — ceratobranchiale, нижній зябровий — hypobranciale, розташовані відносно один одного подібно тому, як у сучасних акулових.
Активізація розширення зябрової кишки сприяла всмоктуванню більших харчових об'єктів, а збільшення ступеня стискання її порожнини до торкання її вентральної стінки до дорсальної визначило зміну ковтання їжі. На зміну дії миготливого епітелію прийшло проштовхування їжі шляхом зміщення вентральної стінки відносно дорсальної. Це зумовило проковтування харчових шматків набагато більших розмірів. Збільшення розмірів здобичі визначило необхідність її схоплювання і втримання при проштовхуванні у стравохід. Цю функцію прийняла на себе вісцеральна дуга, яка перетворилася в щелепи. Дві перші дуги, розташовуючись поблизу ротового кільця,^ не могли забезпечити достатнього стискання. Вони стали допо-? міжними органами щік. Верхній кінець 3-ї дуги придбав фіксацію на нейрокранії. Нижні кінці правої і лівої третіх дуг з'єдналися t між собою. Таким чином, виник протостильний щелепний апарат (рис. 148)

Рис. 148. Послідовні стадії еволюції щелепної та гіоїдної дуг: І — початкова стадія; II — третя вісцеральна дуга перетворилася у щелепи; III — четверта вісцеральна дуга перетворилася у гіоїд
Щелепний апарат, що представлений тільки однією дугою був притаманний рибам класу акантоди. Ці риби, очевидно, не витримали конкуренції з рибами, у яких розвинувся підвісок щелеп до черепа за допомогою 4-ї вісцеральної дуги, що перетворилася в під'язикову дугу. Цим шляхом розвитку щелепного апарата пішли всі сучасні риби.
Як і в щелепній дузі, так і в гіоїдній дузі відбулася редукція самого верхнього членика. Ця дуга представлена тільки трьома члениками: верхнім під'язиковим epihyale, що позначається hyomandibulare і ceratohyale з hypohyale або власне гіоїдом. Перший фіксує щелепний суглоб до слухової ділянки нейрокранія, а гіоїд — відіграє роль другої нижньої щелепи в акул і вісцерального апарата у вищих риб.
Скелет вісцерального апарата сучасних акул цілком хрящовий і представлений щелепною, гіоїдною і зябровими дугами. Спереду від щелепної дуги в основі щік лежать губні хрящі, рудименти 1-ї і 2-ї вісцеральних дуг. Щелепна дуга складається з верхньої щелепи, що називається піднебінно-квадратним (під'язиково-нижньощелепним) хрящем, і нижньої щелепи, або меккелевого хряща. Гіоїдна дуга представлена гіомандибулярним хрящем та гіоїдом. За типом фіксації піднебінно-квадратного хряща до неврального черепа в хребетних розрізняють амфістильний, гіостильний і аутостильний череп.
У амфістильному черепі пінебінно-квадратний хрящ фіксується до нейрокранія в трьох пунктах: в орбітальній ділянці орбітальним відростком або зв'язкою, до слухової ділянки і зв'язками до гіоїдної дуги. Цей тип черепа притаманний примітивним акулам і кістковим ганоїдам.
У гіостильному черепі втрачається зв'язок піднебінно-квадратного хряща зі слуховою ділянкою, а при значному виражені гіостилії — і з орбітальною ділянкою.
Піднебінно-квадратний хрящ у цьому випадку виявляється підвішеним до нейрокранія тільки на гіоманди-булярі. Постильний тип черепа притаманний деяким акулам і осетровим (рис. 149-150).
В аутостильному черепі гіомандибуляр не бере участі у прикріпленні піднебінно-квадратного хряща, який більш-менш тісно зростається з нейрокранієм у нюховій і слуховій ділянках або навіть на всьому протязі (наземні хребетні, рис. 149). Зябрових дуг у акул 5, рідше 6 або 7. Верхні їх членики не зв'язані з нейрокранієм, нижні ж приєднуються до копулярної пластинки, яка утворена базібранхі-аліями, що зрослися, але відокремлені від базігіале. Зяброві перегородки гіоїдної і зябрових дуг продовжуються у кришки відповідних вісцеральних щілин.
У кісткових ганоїдів і кісткових риб прогресивний розвиток щелепного апарата виражається в окостенінні каудального відділу піднебінно-квадратного і меккелевого хрящів, у розвитку на передньому відділі кожного з них ряду покривних кісток і в появі вторинних щелепних кісток.
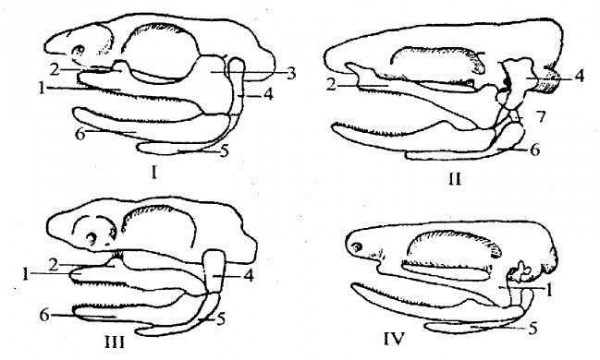
Рис. 149. Типи приєднання щелепного апарата до нейрокранію: І — амфі-стильний череп акули; II — амфістильний череп кісткового ганоїда; III — гіостильний череп акули; IV — аутостильний череп наземного хребетного; 1 — піднебінно-квадратний хрящ; 2 — піднебінноносовий відросток; 3 — вушний відросток; 4 — гіомандибула; 5 — гіоїд, 6 — мекеллів хрящ; 7 — інтергіале,

Рис. 150. Амфістильний череп плащеносної акули (за Аллісом Е. P., 1897 p.): 1 — орбітальний відросток; 2 — піднебінно-квадратний хрящ; 3 — суглоб гіо-мандибули; 4 — гіомандибула; 5 — гіоїд, 6 — мекеллів хрящ; 7 — губні хрящі
За рахунок окостеніння каудального відділу піднебінно-квадратного хряща виникають квадратна і верхня крилоподібна кістки (os quadratum et os metapterygoideum). Спереду від них на цьому хрящі з'являються покривні зовнішня і внутрішня крилоподібні кістки (оssa ecto- et endopterygoidea), а на його передньому кінці змішаного походження піднебінна кістка (os dermo-autopalatinum). Характерно для костистих риб також злиття кісток, що розвивається на піднебінно-квадратному хрящі з гіомандибуля-ром, що костеніє в єдиний функціональний комплекс — гіопалятин-ний апарат. За рахунок окостеніння заднього кінця меккелевого хряща виникає суглобова кіскта (os articulare), що утворює із квадратною кісткою щелепний суглоб. Спереду від неї на цьому хрящі з'являється ряд покривних кісток. Найбільша серед них — зубна кістка (os dentale) — лежить на його передньому відділі. Каудально від неї лежить пластинчаста кістка (os spleniale), кутова (os angulare), надкутова (os supmangulare).
Вториннощелепні кістки, а саме верхньощелепна і передверхньо-щелепна (os maxillare et os praemaxillare) розвиваються як покривні кістки першої і другої передщелепних дуг. Вони не зв'язані з кістками піднебінно-квадратного комплексу і у костистих риб мають досить різноманітну будову, що обумовлено характером харчування.
У всіх костистих риб виникає вторинна чотиричленні сть гіоїдної дуги, це відбувається шляхом диференціації гіомандібули на власне huomadibulare і sympleeticum, а гіоїда на interhyale і власне гіоїд. Характерна для кісткових риб загальна копулярная пластинка, до якої фіксується дистальний кінець гіоїда і всіх зябрових дуг. Прогресивно розвивається при цьому і перегородка гіоїдної дуги, що перетворюється в загальну зяброву кришку. Периферичні відділи зябрових перегородок редукуються. Про збільшення ролі вісцерального апарата в проштовхуванні їжі свідчить розвиток на зябрових дугах зубних площадок.
Кистеперим рибам властиві специфічні особливості будови вісцерального апарата. Так у відомих представників із ряду Osteolepiformes піднебінноквад-ратний блок являє собою єдине окостєніння, у якому умовно розрізняють три відділи, що можна порівняти з quadratum, metap-terygoideum (epipterygoideum) і autopalatinum сучасних целакан-тів. На палятоквадратум остеоле-піформів розвинулися також три покривні кістки: розташована спереду — dermopalatinum, за нею — ectopterygoideum і медіально велика — entopterygoideum. Латерально ці кістки примикають до верхньощелепної і перед-щелепної кісток, що у свою чергу тісно зв'язані з нижнім краєм етмосфеноїдного щита (рис. 151). Квадратний відділ піднебінно-квадратного комплексу сильно витягнутий каудально, через що щелепний суглоб розташовується за рівнем потилиці.
Піднебінно-квадратний комплекс приєднується до неврального черепа в рипідистій у решітчастій, клиноподібній і вушній ділянках. До етмоїду аутопалатінум приєднується медіальним і латеральним зчленуванням. У клиноподібній ділянці піднебінно-квадратний комплекс базальним відростком з'єднувався з базіптерігоїдним відростком нейрокранію, а висхідним відростком — із задньою бічною стінкою етмосфеноїда (ргос. antoticus, рис. 151). Зв'язок із вушною ділянкою непостійний, він підсилюється у прогресивних форм шляхом розвитку вушного піднебінно-квадратного відростка.
Ззаду до піднебінно-квадратного комплексу тісно приєднується під'язиково-нижньощелепна кістка. Цей елемент заповнює нижню частину спіракуло-гіомандибулярної ніші заднього краю піднебінно-квадратного комплексу (рис. 152). Проксимально гіомандібула двома зчленуваннями, розташованими по косій лінії зверху вниз і вперед, приєднується до слухової ділянки нейрокранію. Дистальний кінець гіомандібули через стілогіоїд з'єднувався з гіоїдом. Тісне з'єднання піднебінно-квадратного комплексу з гіомандібулою свідчить про їхню функціональну єдність, до подібного гіопалятинного апарата костистих риб. У рипідістій рухи цього комплексу також тісно пов'язані із змінами положення етмосфеноїдного блоку. Опускання переднього кінця останнього було пов'язане з приведенням піднебін-но-квадратно-гіомандибулярного комплексу, а його піднімання — з відведенням цього комплексу.

Рис. 152. Череп еустеноптерона з піднебінною дугою і підвіском: 1 — переднє та 2 — заднє окостеніння піднебінно-квадратного хряща, 3 — гіоман-дибула; 4 — ентоптерігоїд; 5 — ектоптерігоїд; 6 — піднебінна кістка

Рис. 153. Кістки нижньої щелепи рипідістії: І — латеральна поверхня, II — медіальна поверхня; 1 — зубна; 2 — надкутова; 3 — кутова; 4 — пластинчасті; 5 — суглобова; 6 — передсуглобова; 7 — вінцева
Нижня щелепа рипідистій складалася з суглобової кістки, яка представляла задній окостенілий кінець меккелевого хряща, що бере участь в утворенні щелепного суглоба і ряду покривних кісток. Найбільшою з них була зубна кістка, яка займала верхню частину зовнішньої поверхні нижньої щелепи, що брала участь в утворенні нижньощелепного симфізу і несла ряд зубів. Нижче неї спереду назад розташовувалися такі кістки: пластинчасті, запластинчасті, кутові і надкутові. На внутрішній поверхні нижньої щелепи лежала довга передсуглобова кістка, а по верхньому краю — три вінцеві кістки (os corvnale), що несли зуби (рис. 153).
Date: 2016-11-17; view: 1986; Нарушение авторских прав