Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Регуляция действия генов
|
|
Регуляция действия генов у прокариот и эукариот различна.
Оперонная система регуляции действия генов у прокариот
Экспрессия генов – процесс передачи от ДНК через РНК к полипептидам. Организация генов у прокариот и эукариот различна. Гены прокариот непрерывны, а гены эукариот имеют мозаичную структуру. Так же для прокариот характерен оперонный тип организации генов.
Выживаемость бактерий зависит от способности переключать метаболизм с одного субстрата на другой. Для такой способности гены бактерий объединены в кластеры таким образом, что, ферменты необходимые для определения пути биосинтеза, кодируются генами, находящимися под общим контролем.
Вся система, включающая структурные гены и элементы, контролирующие их экспрессию называют опероном (единица транскрипции и регуляции у бактерий).
В 1961 году была разработана схема регуляции действия генов на уровне транскрипции на примере lac-оперона E. сoli. Результатом активности оперона является синтез, как правило, родственных по биохимическим функциям адаптивных ферментов (конститутивные постоянно находятся в клетке, а адаптивные появляются в результате изменения условий среды). Конститутивные ферменты у E. сoli утилизируют глюкозу. При замене глюкозы на лактозу становится необходима β-галактозидаза и галактозидпермиаза. Эти ферменты кодируются 3 структурными генами: Z, Y, A. Они в комплексе с промотором и терминатором транскрипции образуют оперон. Так же в схему оперонной регуляции входит ген регулятор, кодирующий белок-репрессор. Ген регулятор не входит в состав оперона. Синтезируемый им репрессор, блокирует транскрипцию оперона в присутствии глюкозы.
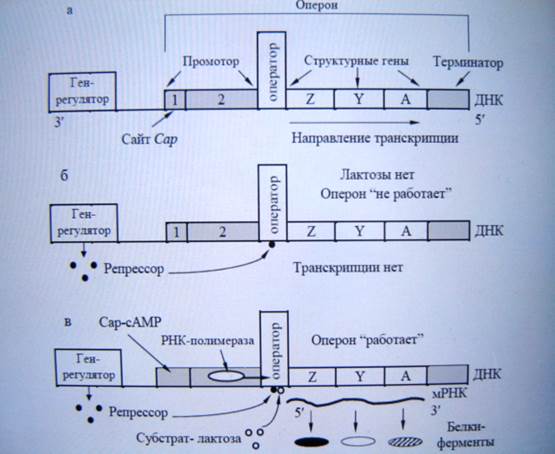
Хотя РНК-полимераза может связаться с промотором, она не перемещается дальше «выключенного» оператора. И гены Z, Y, A не работают. При замене глюкозы на лактозу белок репрессор временно инактивируется под действием субстрата лактозы. Белок репрессор отсоединяется от оперона, что открывает путь молекулам РНК полимеразы для перемещения и транскрипции 3-х генов. Т.е. структурные гены освобождённые от контроля оператора начинают экспрессироваться, продуцируя адаптивные ферменты. Снижение концентрации лактоза служит сигналом для соединения белка репрессора с оператором и прекращение транскрипции lac-оперона.
Удалённость гена-оператора от оперона свидетельствует о том, что репрессор – растворимый в цитоплазме продукт присутствующий в активной форме при наличие в среде глюкозы.
Схема регуляции действия генов lac-оперона у E. сoli называется негативной, т.к. продукт гена регулятора запрещает транскрипцию, т.е. негативно действует на экспрессию.
Позже было установлено, что наряду с негативной системой регуляции lac-оперон находится и по позитивным контролем. Если выращивать на среде глюкоза+лактоза, то утилизация лактозы начнется только после использования всей глюкозы. Транскрипция lac оперона контролируется молекулой эффектором – циклическим AMP (c AMP) и белком активатором Cap-белком. Их присутствие не репрессию, а активацию транскрипции. Вступив в комплекс с c AMP, Cap-белок активируется и только после этого присоединяется к своему сайту на промоторе, многократно усиливая транскрипцию генов lac-оперона. Причем транскрипция lac-генов возможна только при недостатке или отсутствии глюкозы. При избыточном количестве глюкозы комплекс c AMP+Cap-белок не образуется. c AMP получил в бактериях название «сигнал голода».
Основным преимуществом оперонной организации генов у прокариот является координация регулярности активности.
Каскадный тип регуляции у эукариот
Регуляция активности генов эукариот гораздо сложнее и менее изучена. Во-первых у них имеется обособленное ядро и сложно устроенные хромосомы. И, следовательно, транскрипция и трансляция оказались разделены во времени и пространстве. Во-вторых эукариоты (почти все) многоклеточные организмы с чёткой специализацией тканей и органов.
Структурные гены эукариот подразделяются на:
1. гены «домашнего хозяйства». Функционируют во всех клетках организма на всех стадиях (гены кодирующие белки-гистоны, рРНК, тРНК, ДНК полимеразу).
2. «гены роскоши». Функционируют в клетках одной ткани или специфичные для одного типа клеток (гены гемоглобина).
Регуляция действия генов может происходить на любом этапе синтеза белка. ДНК → РНК → белок. Причем механизм регуляции действия генов у эукариот более гибкий. У прокариот по принципу «всё или ничего», а у эукариот от полной экспрессии до полной репрессии. Это достигается за счёт локальной модификации ДНК, изменения внутриклеточных условий.
Регуляция активности генов на уровне транскрипции.
У эукариот имеются специализированные элементы регуляции:
1. Энхансеры – усилители.
2. Сайленсеры – глушители.
Это короткие нуклеотидные последовательности. Их действие связано с изменением структуры хроматина.
Энхансерные последовательности служат в качестве специфических участков связывания особых регуляторных белков, активирующих процесс транскрипции. Они могут располагаться в любой ориентации и на любом расстоянии от промотора.
ДНК между энхансером и промотором образует петлю, в результате чего белки связанные с энхансером, непосредственно взаимодействует с одним из общих факторов транскрипции или с молекулой самой ДНК полимеразы. С помощью белков регуляторов каждый ген специфически включается и выключается.
Согласованная регуляция генов.
У эукариот, гены, контролирующие различные этапы одного и другого пути метаболизма разбросаны по всему геному. Они могут располагаться даже в разных хромосомах. Однако, для них характерно согласованная регуляция действия генов. В ответ на действие различных индукторов, активизируется целая батарея генов. Этому способствует существование гомологичных повторов с 5’ конца у разных генов. Сигналы индукции служат как бы ключами, открывающими замок в различных генах обной группы. Примером согласованной регуляции может быть активация генов теплового шока на тепловой шок.
Гомеобокс и его участие в регуляции генов
В 90 гг. 20 века было обнаружено, что многие гены контролирующие пространственную организацию развития эмбриона, содержат один и тот же сегмент ДНК в виде специфической консервативной последовательности длиной 180 пн, расположенной в 3’ частях генов. Эта последовательность получила название – ГОМЕОБОКС. Гомеобокс кодирует соответствующий ему фрагмент белковой молекулы, длиной 60 АК остатков (гомеодомен). ДНК связывающий домен полипептида относится к регуляторным белкам (Домен – участок полипептида, выполняющий определённую функцию). Т.е. гены, входящие в состав гомеобокса, обладают способностью регулировать активность других генов, путём связывания образующейся при их трансляции АК последовательности с ДНК, включая или выключая при этом определённые гены.
Метелирование ДНК, как способ контроля активности генов.
Метелирование – процесс присоединения –CH3 группы к нуклеотиду, при помощи метилазы. Это обратимая химическая модификация азотистых оснований. В природе широко распространено метилирование цитозина. Каждый сайт метелирования наследуется дочерними ДНК при репликации. В начале 80 гг. была выявлена корреляция между характером экспрессии генов и степени метилирования ДНК. ДНК неактивных генов метилирована сильнее, чем ДНК активных. Мети. ДНК может включать в себя прямое подавление транскрипции путём сворачивания метилированного промотора в недоступную для транскрипции структуру. Степень метилирования ДНК может меняться от различных факторов (возраст, гормональные воздействия, экологические факторы и др.).
Метилирование ДНК – один из основных механизмов эпигенетической изменчивости – изменении дифференциальной экспрессии генов, наследуемые в ряду митотических делений клетки без изменения первичной последовательности нуклеотидов ДНК. Метилирование ДНК изменяет взаимодействие между ДНК и белками, входящих в состав хроматина. И именно это взаимодействие через механизм компактизации – декомпактизации хроматина регулирует экспрессию генов. Только в декомпактизованном состоянии гены, как участники молекулы ДНК становятся доступными ферментам, осуществляющим транскрипцию.
Предполагается, что метилирование ДНК играет важную роль в защите генома от мобильных генетических элементов, а так же в инактивации перенесённых в клетку «чужеродных» генов. До 45 % генома человека занимают вирусные и бактериальные гены, которые, благодаря метилированию молчат. Так же с метилированием связано такое явление как импринтинг – ядерная память – зависимость проявления (экспрессии) гена от того, от кого наследуется данный ген. Т.е. в участках генома, подверженных импринтингу, экспрессируется только один аллель – отца или матери. У человека с феноменом импринтинга связан целый рад наследственных заболеваний.
Регуляция генной активности под действием сигналов внутренней среды.
Примером может служить образование хромосом типа «ламповых щёток» в диплотене мейоза. Мейоз надолго останавливается в профазе 1 мейоза на стадии диплотены. В течении этого времени в ооците происходит интенсивный синтез РНК, ферментов и запасных питательных белковых веществ, необходимых для дальнейшего развития яйца и зародыша. В течении этого периода хромосомы приобретают гигантские размеры и находятся в сильно декомпактизованном состоянии. На более поздних стадиях мейоза хромосомы становятся компактными. На стадии диплотены хромосомы имеют хромомерное строение. Гомологичные хромосомы, соединяясь попарно образуют биваленты. Характеризуются наличием парносимметричных разноразмерных боковых петель, каждая из которых представляет раскрученную хромомеру. На петлях происходит синтез мРНК.
Вытянутые петли и большое количество РНК транскрипт придают хромосомам необычный «мохнатый» вид – щёток. К концу профазы петли снова втягиваются. В образовании петли участвует сразу не весь отрезок ДНК, а в каждый момент только часть этого отрезка. Волна деспирализации протягивается по нему постепенно, пока он не будет полностью транскрибирован.
Регуляция активности генов под действием экстремальных факторов
Механизм такой регуляции, обеспечивается процессами, стремящимися вернуть клетку в прежнее состояние. Важной особенностью подобной регуляции является обратимость. Гены, кодирующие адаптивные (стрессовые) белки, образование которых резко увеличивается под влиянием стрессовых факторов, содержат в составе промоторов дополнительные короткие нуклеотидные последовательности. Например, при тепловом шоке вырабатываются БТШ, быстрое накопление которых в клетке обеспечивает физиологическую адоптацию к изменившимся условиям среды.
Синтез БТШ – это стрессовая программа, включаемая тепловым шоком или многочисленными другими стрессовыми факторами. В ответ на стресс особый полипептид – фактор транскрипции при тепловом шоке связывается с регуляторной зоной генов теплового шока и включает активную транскрипцию данных генов. О значительной роли БТШ жизни клеток, говорит их эволюционный консерватизм (у E. coli, растениях, насекомых, млекопитающих сходны по АК составу). После окончания теплового шока синтез БТШ прекращается и начинается синтез нормальных белков. БТШ выполняют функции шаперонов – белков, способствующих формированию правильной пространственной конфигурации других белков.
Регуляция экспрессии генов под действием гормонов
Гормоны так же играют важную роль в регуляции экспрессии генов. Это половые гормоны, адреналин, гормоны роста, гормоны щитовидной железы и др. Для каждого из них имеются группы компетентных клеток, содержащих специальные рецепторные белки с которыми специфически связываются молекулы гормона. Белки-рецепторы обеспечивают реакцию клеток на воздействия различных гормонов, активируя или подавляя экспрессию определённых генов. Крупные молекулы пептидных гормонов не проникают в клетки и их эффекты, обеспечиваются белками – рецепторами, локализованные на мембранах клеток рецепторов. Образующийся гормон-рецепторный комплекс проникает в ядро и, связываясь со специфическими участками хроматина, переводит его в функционально активное состояние. Следствием этого является транскрипция данного гена, созревание мРНК и транспорт её в цитоплазму и синтез белка.
Каскадная регуляция
Генная активность может регулироваться в процессе онтогенеза. Каскадная регуляция – дифференцированное включение и выключение генов в процессе онтогенеза в различных тканях и органах. Каскадная регуляция происходит на фоне согласованной регуляции в действии многих генов. Т.е. происходит строго упорядоченная во времени и пространстве скоординированная экспрессия сотен тысяч генов.
Немаловажное значение в регуляции действия генов принадлежит и мобильным генетическим элементам. Они могут играть роль мигрирующих промоторов, объединяя структурные гены и регуляторные элементы и настраивая их на общие сигналы регуляции.
Date: 2015-09-24; view: 5596; Нарушение авторских прав