Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Імуногенетика як наука
|
|
Імуногенетика − це комплексна наука, яка вивчає спадкові чинники імунітету, внутрівидову різноманітність і успадкування тканинних антигенів, генетичні і популяційні аспекти взаємин макро- і мікроорганізму та тканинну несумісність.
Методи імуногенетики: використовує методи імунології, молекулярної біології та генетики.
Історія розвитку
Розвиток імуногенетики почався з 1900 року, коли лауреат Нобелівської премії К. Ландштейнер відкрив І-ІІІ групи крові системи АВ0 у людини.
У 1907 році К.Янський відкрив ІУ групу крові системи АВ0 людини.
У 1911 році у людей була знайдена система груп крові МN, яка відрізняється від системи АВО тим, що немає природних антитіл до антигенів М та N.
У 1925 році Бернштейн довів, що групи крові людини системи АВ0 успадковуються.
У 1940 році К. Ландштейнер і Вінер відкрили у людини систему груп крові резус-фактор (Rh).
Розвиток імуногенетики тварин розпочався у 1900 році, коли П. Ерліх та Д. Морген рот відкрили групи крові у кіз. Ініціатором роботи по вивченню індивідуальних особливостей крові в інших сільськогосподарських тварин був М.Ірвін.
Термін „імуногенетика” був запропонований у 1936 році М. Ірвіном.
2. Генетичні системи груп крові тварин і людини
Групи крові людини
У 1900 році К. Ландштейнер у крові людини виявив на еритроцитах крові два природні антигени, названі А і В, а у плазмі – два комплементарні їм природні антитіла a і b.
В імунології антигенами називають сторонні тіла (бактерії, віруси, токсини, білки, полісахариди тощо), які потрапляють в організм тварин. Антигеном може бути будь-яка чужорідна для даного організму молекула з масою понад 5-10 тисяч і достатньо жорсткою структурою. У відповідь на проникнення антигенів в організмі утворюються комплементарні їм антитіла – речовини білкового походження. Останні, взаємодіючи з антигенами, знешкоджують їх.
До 1925 року Бернштейну вдалося з’ясувати, що антигени А і В та антитіла a і b у людини успадковуються і контролюються одним локусом хромосоми.
Як виявилось пізніше, у крові людини є більше ніж 100-200 природних антигенів та антитіл, які контролюються 14-20 локусами. Антигени та антитіла, які контролюються одним локусом, об’єднали в одну систему груп крові. Зокрема, антигени А і В та антитіла a і b ввійшли до системи АВ0.
Генетична система АВ0 груп крові людини
| 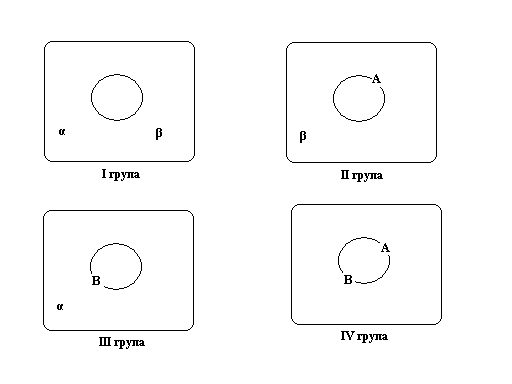 Генетична система АВ0 груп крові людини (у людей з першою групою крові на поверхні еритроцитів знаходяться неактивні антигени 0, а у плазмі крові є антитіла a і b; з другою – антигени А, антитіла b; з третьою – антигени В і антитіла a; з четвертою – антигени А і В, антитіл немає)
Генетична система АВ0 груп крові людини (у людей з першою групою крові на поверхні еритроцитів знаходяться неактивні антигени 0, а у плазмі крові є антитіла a і b; з другою – антигени А, антитіла b; з третьою – антигени В і антитіла a; з четвертою – антигени А і В, антитіл немає)
|
Якщо, наприклад, людині з другою групою крові (антигени А та антитіла b), перелити кров третьої групи (антигени В і антитіла a), то це призведе до взаємодії комплементарних антитіл і антигенів (А з a та В з b), результатом якої є аглютинація, або склеювання еритроцитів. Це, у свою чергу, зумовить закупорку судин і смерть. Тому, у крові однієї людини ніколи не зустрічаються одночасно антигени А з антитілами α, а антигени В з антитілами β.
Генетична система груп крові
Генетична система груп крові − це серія еритроцитарних антигенів та антитіл, які контролюються одним локусом хромосоми (тому, у людини є не 4, а щонайменше 14 генетичних груп крові. Вище згадана генетична система груп крові АВ0 має значення при переливанні крові. Саме всередині цієї системи умовно кров поділяють на чотири групи, про які ми знаємо. Інша, нам знайома, система крові людини, яка називається резус-фактор (Rh+), теж викликає ускладнення при переливанні. І як було згадано раніше, таких систем у людини є щонайменше 14).
Еритроцитарні антигени, аглютиногени, або групові фактори − це антигени, відкриті К. Ландштейнером на еритроцитах, що представляють собою складні біополімерні макромолекули, які містять 85% вуглеводів і 15% білків (антигенна специфічність обумовлена вуглеводним компонентом).
Аглютиніни − це антитіла, відкриті К. Ландштейнером у плазмі.
Групи крові у сільськогосподарських тварин
На теперішній час генетичні системи груп крові вивчені в основних видів сільськогосподарських тварин та деяких диких. Наприклад, у великої рогатої худоби їх відомо 12. Це означає, що за їх успадкування відповідає 12 локусів хромосом. Кожна із систем включає від одного до десятків антигенів. Зокрема, система R–S включає два антигени (R΄, S΄), які детермінуються геном, що може перебувати лише у двох альтернативних станах. Система В об’єднує більше 30 антигенів, які визначаються серією більше 30 множинних алелей.
Генетичні системи груп крові у сільськогосподарських тварин
| Генетичні системи груп крові
великої рогатої худоби
|
У великої рогатої худоби на даний час виявлено 12 систем груп крові: А, В, С, F, J, L, M, S, Z, R-S, T΄, N, які контролюють синтез більше 100 антигенів. Найскладнішою є система В, яка включає більше 30 антигенів, і в різних комбінаціях утворює більше 500 алелей. Приблизно 10 антигенів В-системи успадковуються єдиним комплексом, наприклад, ВBGK, BBG та ін.
У свиней виявлено 17 генетичних систем груп крові (A, B, C, D, E, F, G, H, I, J, K, L, M, N, O, P, Q), які контролюють 83 еритроцитарні антигени. Найскладніші: Е (16 антигенів), L (13 антигенів) і М (11 антигенів). Інші включають 2-6 антигенів. Антигени у всіх системах позначаються великою літерою системи з буквеними індексами: Fa, Fb, Fc, Fd або Ba, Bb.
Групи крові коней включають більше 9 систем (A, C, D, K, Q, Fc, T, U, Sо), які контролюють 20 антигенних факторів. Системи C, K, U, Fc включають по дві алелі, інші – більше двох. Найскладнішою є система D, яка включає 13 антигенів, які утворюють більше 30 феногруп. Антигени у всіх системах позначаються великою літерою системи з індексами: Da, Db, Dc, Dd.
У курей на теперішній час відкрито 14 систем (A, B, C, D, E, H, J, Y, K, Z, N, P, R, Vh), які контролюють 95 антигенів. Найскладніша система В включає 35 антигенів. Встановлено, що антигени систем А та В представляють собою складні комплекси, які включають багато антигенних детермінант. Кожен антигенний комплекс успадковується як самостійна одиниця, що не має собі подібних в інших популяціях. Це створює певні труднощі при уніфікації груп крові курей. Антигени позначаються буквою відповідної генетичної системи з підстрочною цифрою – В4, а при позначенні алелей до великої літери добавляють надстрочну цифру – В4.
Групи крові овець менше вивчені, ніж інших тварин. До теперішнього часу виявлено 16 генетичних систем (A, B, C, D, J, M, R, X-Z, Con, F30, F4, Hel, Y, T, V, PV), які контролюють 39 антигенів.
У кіз ідентифіковано 5 систем груп крові (B, C, M, R, F30).
Вивчення генетичних систем груп крові у основних видів сільськогосподарських не завершене. Саме тому у літературі є суперечливі дані. Так, за даними інших авторів, у свиней є не 17, а 14 систем, у овець не 16, а 7, у коней не більше 9, а 8 тощо.
Тип крові
Тип крові − це сума груп крові всіх генетичних систем особини.
Кількість можливих комбінацій груп крові з урахуванням всіх систем настільки значна, що, наприклад, у великої рогатої худоби вона значно перевищує загальне поголів'я у всьому світі. Цим і пояснюється індивідуальна різноманітність тварин всередині виду за типом крові, оскільки кожній особині відповідає своя комбінація груп крові. Тільки однояйцеві близнюки мають повністю ідентичні типи крові.
Міжнародна номенклатура антигенів і систем крові
До теперішнього часу міжнародна номенклатура антигенів і систем крові не уніфікована і для різних видів тварин має свої особливості. Щоправда з 1928 року як групи крові так і антигени прийнято позначати літерами латинського алфавіту – А, В, С і т. д. Якщо антигенів багато, а літер алфавіту не вистачає, їх записують зі штрихами – А', В', С' та з індексами – А1, А2 і т. д. Необхідно пам'ятати, що А та А' – це різні антигени, не пов'язані генетично та імунологічно один з одним. Іноді антигени позначають двома літерами: великою та малою, наприклад, Еа, Ее тощо.
Феногрупи − групи антигенів деяких систем з визначеною комбінацією включених антигенів (у великої рогатої худоби групи крові В, G та К можуть зустрічатися в комбінаціях: тільки В, тільки G, ВG, ВGК. Можлива і повна відсутність всіх наведених факторів. Для спрощення записів феногрупи кодують. Наприклад, феногрупу BGKO2Y1A'B'E'K'O'Y' позначають В28. Генотипи за групами крові записують так: ВBGВBGK, або ВBG/ВBGK).
Проста система груп крові містить один-два антигени та має лише дві алелі (у великої рогатої худоби це система N).
Складна система груп крові (поліалельна) включає три антигени й більше, які контролюються серією множинних алелей (у великої рогатої худоби це системи В та С).
У закритій системі за антигенами еритроцитів можна визначити генотип тварини за цією системою.
У відкритій системі визначити генотип тварини за цією системою можна лише у деяких гомозигот.
Успадкування груп крові
1. Гени, які визначають антигени систем груп крові, локалізовані в аутосомах.
2. Гени, які кодують еритроцитарні антигени однієї генетичної системи груп крові, можуть бути представлені двома алелями, або утворювати серію множинних алелей.
3. Генотип за певною системою крові може бути гомо- (А/А) чи гетерозиготним (А/В). Гомозиготних особин виявляють за гемолітичними тестами (відсутність реакції), гетерозиготних – сімейним аналізом.
4. У деяких системах є алелі, які контролюють відсутність антигенів. Їх називаються рецесивними, нульовими, або негативними і позначають знаком віднімання (–), або малою буквою латинського алфавіту (а, b тощо).
5. Антигени різних генетичних систем можуть успадковуватись незалежно один від одного та зчеплено.
6. Зчеплене успадкування антигенів різних генетичних систем пояснюється тісним зчепленням локусів, які контролюють ці антигени. Антигени, які успадковуються зчеплено, утворюють феногрупи. Такий тип успадкування характерний для складних систем. Приблизно 10 антигенів В-системи (див. табл.) великої рогатої худоби успадковуються єдиним комплексом, наприклад, ВBGK, BBG та ін.
7. Більшість алелей генетичних систем груп крові успадковуються за типом кодомінування, тобто у гетерозиготних особин обидва гени виявляють свою дію повною мірою незалежно один від одного. За таким типом, наприклад, у великої рогатої худоби успадковуються алелі F-системи:
| P | ♀V/V | x | ♂ F/F |
| антиген V | антиген F | ||
| G | V | F | |
| F1 | V/F антигени V та F |
8. Іноді зустрічаються рецесивні алелі, зокрема в системі АB0 у людини, де алель 0 рецесивна щодо алелей А і В, хоча останні успадковуються за кодомінантним типом:
Ø алель А домінантна, 0 – рецесивна:
| P | ♀0/0 | x | ♂ А/А |
| антиген відсутній | антиген А | ||
| G | А | ||
| F1 | 0/А антиген А |
Ø алель В домінантна, 0 – рецесивна:
| P | ♀0/0 | х | ♂ В/В |
| антиген відсутній | антиген В | ||
| G | В | ||
| F1 | 0/В антиген В |
Ø алелі А та В кодомінантні:
| P | ♀А/А | x | ♂ В/В |
| антиген А | антиген В | ||
| G | А | В | |
| F1 | А/В антигени А та В |
9. Кожна особина успадковує по одній з двох алелей від матері і батька в кожній генетичній системі.
10. Особина з антигенами, які не виявлені хоча б у одного з батьків, не може бути нащадком даної батьківської пари.
11. Гомозиготна особина за одним антигеном, наприклад FF/F, не може бути нащадком гомозиготної особини з протилежним антигеном, наприклад FV/V.
Успадкування груп крові у людини
Система АВ0визначається одним аутосомним геном, який існує в трьох алельних станах – і°, ІА, ІВ. Алелі ІА та ІВ домінують над і° і виступають кодомінантними по відношенню один до одного в гетерозигот, тобто кожна з цих алелей проявляється у фенотипі.
| Система MN визначається одним аутосомним геном L, який існує у двох кодомінантних алельних станах LM и LN. Кожен з алелів відповідає за вироблення відповідного еритроцитарного фактора: LM – М (група крові М), LN – N (група крові N). У гетерозигот за геном L на еритроцитах виявляються обидва фактори – група крові MN.
|
Антигени в системах MN та АВ0 успадковуються за кодомінантним типом:
Успадкування групи кровіАВ0 у людини
| P | ♀АА х ♂ВВ | |
| ІІ група антиген А | ІІІ група антиген В | |
| G | А | В |
| F1 | АВ | |
| 100% ІV група (антигени А та В) |
Усі групи крові використовую у сімейно-генетичному аналізі для встановлення ступеня рідні. При цьому виходять з положення, що в нащадка не може бути гена, якого немає в батьків, та в той же час нащадок може не успадкувати гена, присутнього в батьків.
3. Імуногенетична несумісність матері і плода
Система резус-фактор (Rh) та резус-конфліктна ситуація між матір’ю і плодом через резус-несумісність
Система резус (Rh) контролюється аутосомним геном із двома алельними станами, резус-позитивність (Rh+) домінує над резус-негативністю (Rh-).
| Назва групи крові | Антиген | Генотип особини |
| Rh+ (резус-позитивна) | Rh+ | Rh+ Rh+, Rh+ Rh- |
| Rh- (резус-негативна) | Rh- | Rh-Rh- |
Одним а антигенів є резус-фактор, який одержав таку назву тому, що вперше був відкритий у одного з видів мавп – Макаки резус. Приблизно 85% людей на еритроцитах мають резус-антиген. Їх називають резус-позитивними (в серології позначають Rh+), інші 15% його не мають, вони резус-негативні (Rh–).
У 1940 році Лєві відкрив у людини гемолітичну хворобу новонароджених і з’ясували, що вона зумовлена імуногенетичною несумісністю матері і плода через резус-антиген.
Схема розвитку гемолітичної хвороби новонароджених у людини
| Ø у шлюбі між резус-негативною (Rh-Rh-) жінкою і гомозиготним резус-позитивним (Rh+Rh+) чоловіком усі діти будуть гетерозиготні (Rh+Rh-) резус-позитивні; Ø при першій вагітності еритроцити резус-позитивного плода з резус-антигенами проникають через плаценту в організм матері і викликають у ньому утворення специфічних антитіл проти резус-антигенів, які накопичуються в організмі; Ø у послідуючі вагітності після першої благополучної антитіла проникають через плаценту у кров’яне русло ембріона і гемолізують його зрілі еритроцити, що призводить до ембріонального еритробластозу (переважання у крові незрілих еритроцитів), яке найчастіше закінчується смертю. Попередити смерть новонародженого може лише повна заміна крові. Резус-негативна мати, яка виношує резус-позитивний плід, часто страждає на різного ступеня інтоксикації, але це не призводить до абортів. |
|
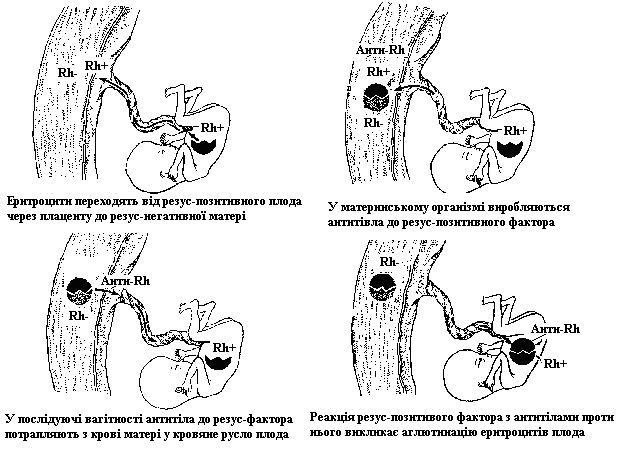
Імовірність виникнення резус-конфліктної ситуації між матір’ю і плодом:
1. Якщо батько гомозиготний резус позитивний, а мати резус-негативні усі діти будуть страждати на еритробластоз.
2. Якщо батько гетерозиготний (Rh+ Rh-) за резус-позитивним фактором, то у середньому імовірно лише половина дітей від таких шлюбів буде страждати на еритробластоз, а інші будуть здоровими. Дуже часто у таких шлюбах усі діти народжуються здоровими, бо успадковують від матері і батька рецесивні алелі.
2. У резус-негативних батьків, усі діти резус-негативні.
3. У гомозиготної чи гетерозиготної резус-позитивної жінки усі діти народжуються здоровими, незалежно від того, який фактор вони успадкували чи який генотип має її чоловік.
Попередження смерті новонародженого від ембріонального еритробластозу: діагностують антитіла у крові матері, які дозволяють прийняти правильні рішення про доцільність кесаревого розтину, або негайного переливання новонародженому резус-позитивної крові (протягом перших 9 годин після народження), щоб встигнути звільнити організм дитини від зруйнованих резус-позитивних еритроцитів і шкідливих продуктів їх розпаду.
Резус-антитіла можуть з’являтися у резус-негативної жінки і без будь-якої вагітності, якщо їй перелити кров від резус-позитивних донорів. Переливання резус-позитивної крові резус-негативним чоловікам теж призводить до утворення в них резус-антитіл. У обох випадках відбувається аглютинація, або склеювання еритроцитів, що, у свою чергу, зумовлює закупорку судин і смерть.
Гемолітична хвороба новонароджених у сільськогосподарських тварин
Гемолітична хвороба новонароджених зустрічається у сільськогосподарських тварин (коней, свиней). Її генетична природа менше вивчена, ніж у людини, але є повне право вважати, що її причиною є також імунологічна несумісність матері і плода.
На відміну від людини, плацента коней та свиней непроникна для антитіл, але вони є у молозиві. Поросята і лошата народжують здоровими, але після ссання матері в перші 12-48 годин у них спостерігаються патологічні зміни у вигляді жовтяниці склери очей (тому цю хворобу ще називають жовтяницею новонароджених), загальної слабкості, збільшення частоти дихання та зниження кількості еритроцитів. Молодняк у таких випадках гине протягом декількох днів.
Для утворення в організмі цих тварин антитіл у кількості, токсичній для новонароджених, однієї вагітності недостатньо. Тільки після другої вагітності від таких імунонесумісних самців титр антигенів у самки досягає рівня, згубного для еритроцитів новонародженого. Тварини від таких самок можуть бути збережені тільки шляхом термінового переливання крові і випоювання їм у перші 24-36 годин молозива інших самок. У цей час молозиво матері здоюють. Через 36 годин шлунково-кишковий тракт новонароджених втрачає здатність засвоювати антитіла такого молозива і їх можна перевести до рідних матерів. Однак, краще за все, зарання визначити титр антитіл у матері (по відношенню до крові самця, з яким її спарювали) і повністю виключити вживання новонародженими молозива матері. У даному випадку можна обійтись без переливання крові.
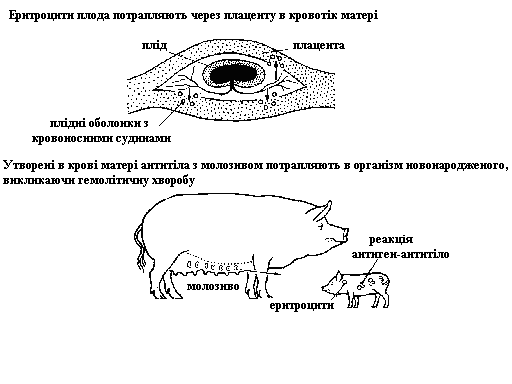
Схема розвитку гемолітичної хвороби новонароджених у свиней
4. Біохімічний поліморфізм білків
Білки – високомолекулярні біополімери, мономерами яких є амінокислоти, які виконують важливі функції в організмі тварин. Зокрема:
Ø трансферин (Тf) – металопротеїн, який зв'язує залізо плазми і переносить у кістковий мозок, де воно використовується для кровотворення;
Ø церулоплазмін (Ср) – металопротеїн, який відіграє основну роль в обміні міді в організмі;
Ø амілаза (Аm) – фермент, який прискорює реакцію гідролізу дегідрогенази в організмі.
Поліморфізм білків
Генетичні типи білків − це фракції одного виду білка у біологічних рідинах (крові, плазмі, м’ясі тощо), які дещо відрізняються за будовою. Типи ферментів, які також мають білкову природу, називаються ізоферментами.
Вперше різні типи білка виявив Л.Полінг, який з’ясував, що у людей гемоглобін буває двох типів – нормальний і мутантний, які відрізняються тільки за однією з 287 амінокислот:
Гемоглобін А – пролін-глютамін-глютамін-лізин
Гемоглобін S – пролін-валін-глютамін-лізин.
Метод виявлення біохімічного поліморфізму білків: метод зонального електрофорезу.
Заснований на розділенні типів білків, які знаходяться у спеціальному (буферному) розчині, під дією електричного струму. При відповідних значеннях рН та іонній силі розчину типи білків переміщуються до анода з різною швидкістю, яка залежить від величини їх заряду. Це явище називають розгонкою. Розганяти білки можна на фільтрувальному папері, крохмальному, агар-агаровому, поліакриламідному гелі тощо.
З удосконаленням у 1955 році Смітісом методу зонального електрофорезу вдалося виявити більше як 150 поліморфних систем білків крові, молока, м’язів, яєць, сперми тощо.
Поліморфна система білків − це система генетичних типів певного білка.
Поліморфні системи білків крові і молока
сільськогосподарських тварин
| У сироватці крові тварин виявлено поліморфізм гемоглобіну, преальбуміну, постальбуміну, гаптоглобуліну, трансферину, церулоплазміну, амілази, лужної фосфатази і карбоангідрази, у сироватці молока – b-лактоальбуміну і b-казеїну. Кількість поліморфних систем поки що невідома, однак вважають, що у популяціях багатьох видів тварин вона досягає 25-50%. Наприклад, у великої рогатої худоби виявлено 10 типів гемоглобіну, у овець – 5, коней – 2. Типів трансферину у великої рогатої худоби є 12, у коней – 10, свиней – 5, овець – 13. |
Поліморфізм білків − це одночасна присутність у популяції двох або більше генетичних типів певного білка.
Номенклатура поліморфних типів білків
За міжнародною номенклатурою поліморфні типи білків прийнято позначати буквеною символікою – TfА, TfВ, TfС, TfD, TfЕ.
Успадкування поліморфних типів білків
Як і групи крові тварин, поліморфні типи білків детермінуються генами, зберігаються протягом життя особини і не піддаються впливу факторів зовнішнього середовища.
З особливостей успадкування поліморфних типів білків відомо, що:
1. Гени, які визначають типи білків і ферментів, локалізовані в аутосомах.
2. Типи білків різних поліморфних систем успадковуються незалежно один від одного.
3. Гени, які кодують типи білків і ферментів однієї поліморфної системи, можуть бути представлені двома алелями, або утворювати серію множинних алелей. Наприклад, у великої рогатої худоби трансферин представлений 12-ма множинними алелями: TfА, TfВ, TfС, TfD, TfЕ тощо.
4. Особини за типами білка чи ферменту можуть бути гомо- (TfА/TfА) і гетерозиготними (TfА/TfВ). У гетерозиготних особин обидва гени виявляють свою дію повною мірою незалежно один від одного, тобто успадковуються кодомінантно:
| P | ♀ TfА/TfА | x | ♂ TfВ/TfВ |
| трансферин А | трансферин В | ||
| G | TfА | TfВ | |
| F1 | TfА/TfВ трансферин А та В |
5. Кожна особина успадковує по одній з двох алелей від матері і батька в кожній поліморфній системі.
6. Особина з типами білків чи ферментів, які не виявлені хоча б у одного з батьків, не може бути нащадком даної батьківської пари.
7. Гомозиготна особина за одним типом білка чи ферменту, наприклад TfА/TfА, не може бути нащадком гомозиготної особини з протилежним типом білка чи ферменту, наприклад TfВ/TfВ.
5. Практичне використання досягнень імуногенетики у тваринництві
Досягнення у вивченні груп крові та поліморфних систем білків і ферментів широко використовуються у тваринництві для:
Ø встановлення і уточнення походження тварин;
Ø визначення зиготності близнят;
Ø генетичного маркування селекційно цінних тварин та прогнозування продуктивності і стійкості проти захворювань їхніх нащадків;
Ø встановлення сумісності батьківських пар при чистопородному розведенні;
Ø прискорення оцінки плідників за якістю нащадків;
Ø побудови генетичних карт хромосом;
Ø визначення фримартинізму у теличок;
Ø виявлення антигенної несумісності організму матері і плода;
Ø вивчення геногеографії видів і порід тварин та генетичних процесів у популяціях;
Ø вивчення причин і динаміки генотипової мінливості та генетичних ресурсів;
Ø виведення інбредних гомозиготних плідників тощо.
Генетичне маркування селекційно цінних тварин та прогнозування продуктивності і стійкості проти захворювань їхніх нащадків
Вивчення груп крові тварин показує, що між ними і господарсько-корисними ознаками та стійкістю до хвороб є корелятивні зв’язки. Тому зараз деякі алелі певних систем груп крові використовують для прогнозування стійкості тварин до хвороб, продуктивності тварин і племінної цінності самців-плідників. Такі алелі називають генетичними маркерами.
Наприклад, при вивченні можливостей використання кореляції груп крові з продуктивністю корів чорно-рябої породи виявлено, що корови, які успадкували від родоначальника лінії бугая Лукавогого алель G3Y3 В-системи давали (табл.) на 720-1111 кг більше молока, ніж корови, які одержали алель BG3O3Y2B. Позитивний зв’язок між групами крові і продуктивністю, встановлена в одних стадах, не підтверджується в інших.
Продуктивність нащадків бугая Лукавого 183
| Ступінь рідні | Номер групи | Алелі, успадковані від родоначальника | Кількість особин (n) | Надій, кг (M±m) |
| Онучки | BG3O3Y2B | 5544±157 | ||
| Онучки | G3Y3 | 6264±159 | ||
| Правнучки | G3Y3 | 6810±292 | ||
| Правнучки | BG3O3Y2B | 5699±194 |
р≤0,01
Найбільш яскравим доказом взаємозв’язку груп крові з резистентністю тварин є кореляція алелі В21 В-системи курей зі стійкістю до хвороби Марека у курей (табл.). Серед особин, гомозиготних за цією алеллю, смертність майже у 2 рази менша, ніж серед гомозиготних за алеллю В19.
Зв’язок груп крові з резистентністю курей до хвороби Марека
| Генотипи | Відхід курчат, % | |||
| Заражені штучно | Заражені природно | |||
| M±m | n | M±m | n | |
| B19/B19 | 12,4±1,20 | 20 100 | 29,0±2,60 | 20 138 |
| Б21/Б21 | 6,5±1,40 | 19 679 | 16,0±2,07 | 19 639 |
р≤0,001
Однак для більшості хвороб тварин поки що відсутні надійні маркери, за якими можна було б успішно вести селекцію на резистентність проти різних інфекційних захворювань. Це свідчить про те, що у цьому напрямку потрібно продовжувати дослідження.
МЕТОДИЧНІ РЕКОМЕНДАЦІЇ ДО ВИКОНАННЯ ІНДИВІДУАЛЬНОГО ЗАВДАННЯ З БІОМЕТРІЇ
Наука, яка розробляє методи вивчення мінливості ознак у живих організмів, називається біометрією. Математичною основою цієї науки є теорія імовірності. Всі ознаки живих організмів поділяють на якісні і кількісні. Величина ознаки в окремої особини називається варіантою. Великий масив живих організмів, який цікавить дослідника, називається генеральною сукупністю. Вивчення великої кількості організмів – складний і дорогоцінний захід, тому застосовують метод випадкової вибірки. Кількість особин (ознак) у вибірці називається об’ємом вибірки і позначається буквою n. Якщо кількість тварин (ознак) у вибірці до 30, вона вважається малою, якщо більше 30 – великою.
Середні величини
Показник, який характеризує генеральну сукупність за величиною показника, який вивчається, називаються середньою арифметичною. Середня арифметична (М) в малочисельних вибірках (коли n<30) розраховується за формулою:
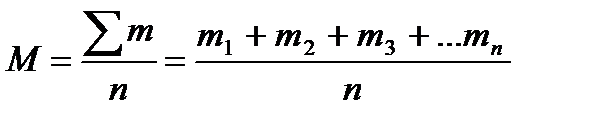 , де
, де
М – середня арифметична; m – величина ознаки в окремої особини (варіанта); ∑х – сума варіант; n – кількість особин (показників) у вибірці (кількість варіант).
Показники різноманітності ознак в сукупності
Ліміти, які характеризують мінімальне (min) і максимальне (max) значення у вибірковій сукупності і вказують на амплітуду варіації, можуть служити показниками різноманітності ознаки в сукупності. Але цих показників недостатньо. Для точного вивчення варіабельності ознаки певної сукупності застосовують такі статистичні параметри як середнє квадратичне відхилення.
Середнє квадратичне відхилення є показником різноманітності ознаки, яка досліджується, для групи, що має визначену середню арифметичну. Наприклад, отримана величина s=0,14 кг вказує, що у середньому відхилення варіант даної ознаки від середньої арифметичної маси поросят складає 0,14 кг. Середнє квадратичне відхилення (s) в малочисельних вибірках (коли n<30) розраховується за формулою:
 , де
, де
s – середнє квадратичне відхилення; m – величина ознаки в окремої особини (варіанта); M –середня арифметична; n – кількість особин (показників) у вибірці (кількість варіант).
Для розрахунку складається робоча таблиця:
| Номер варіанти | Варіанта (m) | Відхилення (m–M) | Квадрат відхилення (m–M)2 |
| n | |||
| ∑(m–M)=0 | ∑(m–M)2 |
У другу графу вписують варіанти (m) (наприклад, жива маса поросят при народженні). Далі варіанти сумують, суму ділять на число варіант і таким чином одержують середню арифметичну (М).
Потім віднімають М (середню арифметичну) від кожної m (варіанти) і різниці (відхилення від середньої) вписують у третю графу. Ці значення можуть бути від’ємними. Для перевірки правильності обчислень підсумовують всі різниці m–M, їхня сума повинна дорівнювати нулю. Далі кожне відхилення підводять до квадрату і вписують квадрати відхилень (m–M)2 у четверту графу. Квадрати відхилень завжди позитивні. Підсумовуючи всі числа четвертої графи, одержують суму квадратів відхилень ∑(m–M)2, яку вписують у підсумок четвертої графи. Далі за даними останнього рядка таблиці обчислюють середньоквадратичне відхилення за вище згаданою формулою.
При вивченні різноманітності ознак, які виражені в різних одиницях виміру (см, кг, % та ін.), і при великій різниці середніх арифметичних величин груп, які порівнюються, сигма не може бути використана. У таких випадках використовують коефіцієнт варіації, який розраховують за формулою:
 , де
, де
С V – коефіцієнт варіації; s – середньоквадратичне відхилення; М – середня арифметична.
Оцінка достовірності одержаних даних
Щоб оцінити на скільки середня арифметична відхиляється від значення окремих варіант, визначають похибку середньої арифметичної (m) за формулою:
Похибку середньої арифметичної (m) для малочисельних вибірок (коли n<30) обраховують за формулою:
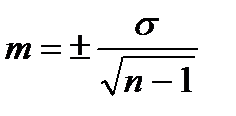 , де
, де
m – похибка середньої арифметичної; σ – середньоквадратичне відхилення; n – кількість варіант у вибірці.
Оцінка достовірності різниці між середніми величинами двох вибірок
У багатьох дослідженнях виникає необхідність порівняти середні арифметичні двох груп тварин (наприклад, середню живу масу тварин дослідної і контрольної груп). Середні арифметичні двох груп, які порівнюються, завжди відрізняються одна від одної. Тому необхідно встановлювати, чи ця різниця достовірна.
Для оцінки вірогідності різниці між середніми величинами різних груп визначають вірогідність різниці (td).
Вірогідність різниці (td) розраховують за формулою:
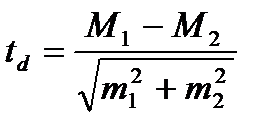 , де
, де
(td) – вірогідність різниці; d = M1 – M2 – різниця між двома середніми (M1 і M2);
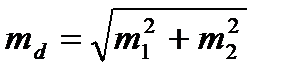 – середня помилка різниці (md).
– середня помилка різниці (md).
Також знаходять число ступенів свободи за формулою:
ν= (n1+n2)–1
Далі за таблицею 1 стандартних значень критерію td для малих вибірок (за Стьюдентом) знаходять достовірність Р.
Наприклад, при td=2,57 імовірність того, що різниця достовірна, складає 0,01 (тобто 99%), а при td=3,2 імовірність досягає 0,001 (99,9%). Якщо ж величина td менша l,96, то різниця між середніми порівнюваних груп не може бути визнана достовірною.
Таблиця 1
Стандартні значення критерію td для малих вибірок (за Стьюдентом)
| Число ступенів свободи ν= (n1+n2)–1 | Достовірність (Р) | ||||
| 0,90 або 0,1 | 0,95 або 0,05 | 0,98 або 0,02 | 0,99 або 0,01 | 0,999 або 0,001 | |
| 6,31 | 12,7 | 31,82 | 63,66 | — | |
| 2,92 | 4,30 | 6,97 | 9,93 | 31,60 | |
| 2,35 | 3,18 | 4,54 | 5,84 | 12,94 | |
| 2,13 | 2,78 | 3,75 | 4,60 | 8,61 | |
| 2,02 | 2,57 | 3,37 | 4,03 | 6,86 | |
| 1,94 | 2,45 | 3,14 | 3,71 | 5,96 | |
| 1,90 | 2,37 | 3,00 | 3,50 | 5,41 | |
| 1,86 | 2,31 | 2,90 | 3,36 | 5,04 | |
| 1,83 | 2,26 | 2,82 | 3,25 | 4,78 | |
| 1,81 | 2,23 | 2,76 | 3,17 | 4,59 | |
| 1,80 | 2,20 | 2,72 | 3,11 | 4,44 | |
| 1,78 | 2,18 | 2,68 | 3,06 | 4,32 | |
| 1,77 | 2,16 | 2,65 | 3,01 | 4,22 | |
| 1,76 | 2,15 | 2,62 | 2,98 | 4,14 | |
| 1,75 | 2,13 | 2,60 | 2,95 | 4,07 | |
| 1,75 | 2,12 | 2,58 | 2,92 | 4,02 | |
| 1,74 | 2,11 | 2,57 | 2,90 | 3,97 | |
| 1,73 | 2,10 | 2,55 | 2,88 | 3,92 | |
| 1,73 | 2,09 | 2,54 | 2,86 | 3,88 | |
| 1,73 | 2,09 | 2,53 | 2,85 | 3,85 | |
| 1,72 | 2,08 | 2,52 | 2,83 | 3,82 | |
| 1,72 | 2,07 | 2,51 | 2,82 | 3,79 | |
| 1,71 | 2,07 | 2,50 | 2,81 | 3,77 | |
| 1,71 | 2,06 | 2,49 | 2,80 | 3,75 | |
| 1,71 | 2,06 | 2,49 | 2,79 | 3,73 | |
| 1,71 | 2,06 | 2,48 | 2,78 | 3,71 | |
| 1,70 | 2,05 | 2,47 | 2,77 | 3,69 | |
| 1,70 | 2,05 | 2,47 | 2,76 | 3,67 | |
| 1,70 | 2,05 | 2,46 | 2,75 | 3,66 | |
| 1,70 | 2,04 | 2,46 | 2,75 | 3,65 | |
| ∞ | 1,64 | 1,96 | 2,33 | 2,58 | 3,29 |
Показники зв‘язку між ознаками (к оефіцієнт фенотипової кореляції)
При одночасному вивченні сукупності тварин за декількома ознаками часто виявляється існування між ними взаємного зв‘язку. Так, крупніші корови мають більш високу молочну продуктивність. Взаємозв‘язок між ознаками в сукупностях живих організмів називається кореляцією.
Вивчення зв‘язку між ознаками має велике значення при вирішенні генетико-селекційних питань. Встановлення фенотипового та генотипового зв‘язку між ознаками дає можливість проводити непряму селекцію за корелюючими ознаками.
За формою кореляція може бути прямолінійною і криволінійною, за напрямком – прямою (позитивною) і оберненою (негативною).
Якщо рівномірним змінам однієї ознаки відповідають рівномірні зміни іншої ознаки при незначних відхиленнях, корелятивний зв'язок прямолінійний. Наприклад, при збільшенні довжини тулуба тварини на 1 см його ширина також збільшується на певну величину.
Якщо зі збільшенням однієї ознаки інша збільшується до певного моменту, а потім зменшується (або навпаки), корелятивний зв'язок криволінійний. Наприклад, зі збільшення віку надій також збільшується до 6-7 отелення, а потім у більшості корів знижується. При криволінійній кореляції зв‘язок спочатку позитивний, потім негативний – тобто зі збільшенням першої ознаки друга, яка з нею корелює, зменшується.
Ступінь зв‘язку між ознаками вимірюється за допомогою коефіцієнту фенотипової кореляції (r) за формулою:
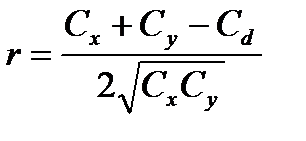
де х і y – значення варіант першої та другої ознак; n – кількість пар ознак, які порівнюються; Cx, Cy, Cd – сума квадратів центральних відхилень, або дисперсії, які обчислюються за формулами:
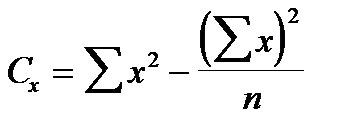 ,
, 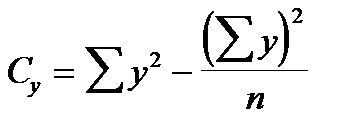 ,
, 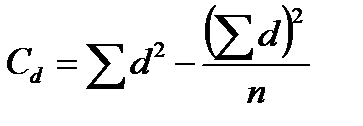 ;
;
d – різниця між першою та другою ознаками.
Властивості коефіцієнту фенотипової кореляції
1. Коефіцієнт кореляції знаходиться в межах від 0 до +1. Якщо r = 0,2-0,3 – зв'язок малий, r = 0,4-0,6 – зв'язок середній, r = 0,7 і більше – зв'язок сильний.
2. Наявність у коефіцієнта кореляції знаку «–» або «+» вказує на напрямок зв‘язку. Якщо коефіцієнт кореляції має знак «+» – зв‘язок між ознаками прямий, тобто зі зміною однієї ознаки інша, корелююча з нею, змінюється у тому ж напрямку. Якщо коефіцієнт кореляції має знак «–» – зв‘язок між ознаками обернений, тобто збільшення однієї ознаки супроводжується зменшенням іншої, і навпаки.
3. Коефіцієнти кореляції виявляють величину і напрямок зв‘язку за умови, що зв‘язок між ознаками наближаються до прямолінійного.
4. Квадрат коефіцієнта кореляції (r2) називають детермінантою. Вона показує частку мінливості ознаки (функції), обумовлену впливом діючого фактора (аргументу).
ПРИКЛАД РОЗВ’ЯЗКУ ТИПОВОГО ЗАВДАННЯ №1:
Завдання 1. Зробити біометричну обробку даних, визначивши середню арифметичну, похибку середньої арифметичної, середнє квадратичне відхилення та коефіцієнт варіації для двох генеральних сукупностей за вмістом пилкових зерен вересу звичайного у меді та оцінити достовірність різниці між середніми величинами двох вибірок.
| Номер зразка | Вміст пилкових зерен вересу звичайного у меді, % | |
| 1 партія відбору (m) | 2 партія відбору (m) | |
Розв’язання:
СЕРЕДНЯ АРИФМЕТИЧНА (М):
Знайдемо середню арифметичну (М1) для партії меду №1:
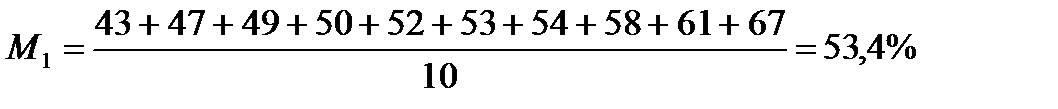
Знайдемо середню арифметичну (М2) для партії меду №2:

СЕРЕДНЄ КВАДРАТИЧНЕ ВІДХИЛЕННЯ (s):
Знайдемо середнє квадратичне відхилення (s) для партії меду №1:
1. Складемо робочу таблицю:
| Номер варіанти | Варіанта (m) | Відхилення (m–M) | Квадрат відхилення (m–M)2 |
| 43–53,4=-10,4 | 108,16 | ||
| 47–53,4=-6,4 | 40,96 | ||
| 49–53,4=-4,4 | 19,36 | ||
| 50–53,4=-3,4 | 11,56 | ||
| 52–53,4=-1,4 | 1,96 | ||
| 53–53,4=-0,4 | 0,16 | ||
| 54–53,4=0,6 | 0,36 | ||
| 58–53,4=4,6 | 21,16 | ||
| 61–53,4=7,6 | 57,76 | ||
| 67–53,4=13,6 | 184,96 | ||
| M=∑m:10=53,4 | ∑(m–M)=0 | ∑(m–M)2=446,4 |
2. У другу графу вписуємо варіанти (m).
3. Варіанти сумуємо, суму (∑) ділимо на число варіант і таким чином одержуємо середню арифметичну (М).
4. Віднімаємо М (середню арифметичну) від кожної m (варіанти) і різниці (відхилення від середньої) вписуємо у третю графу. Ці значення можуть бути від’ємними. Для перевірки правильності обчислень підсумовуємо всі різниці m–M, їхня сума повинна дорівнювати нулю.
5. Кожне відхилення підводимо до квадрату і вписуємо квадрати відхилень (m–M)2 у четверту графу. Квадрати відхилень завжди позитивні. Підсумовуючи всі числа четвертої графи, одержуємо суму квадратів відхилень ∑(m–M)2, яку вписуємо у підсумок четвертої графи.
6. За даними останнього рядка таблиці обчислюємо середньоквадратичне відхилення (s) за вище згаданою формулою: 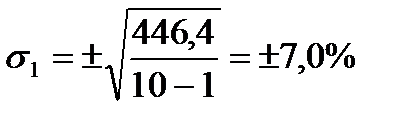
Знайдемо середнє квадратичне відхилення (s) для партії меду №2:
1. Складемо робочу таблицю:
| Номер графи | |||
| Номер варіанти | Варіанта (m) | Відхилення (m–M) | Квадрат відхилення (m–M)2 |
| 50-56,7=-6,7 | 44,89 | ||
| 52-56,7=-4,7 | 22,09 | ||
| 53-56,7=-3,7 | 13,69 | ||
| 49-56,7=-7,7 | 59,29 | ||
| 60-56,7=3,3 | 10,89 | ||
| 63-56,7=6,3 | 39,69 | ||
| 66-56,7=9,3 | 86,49 | ||
| 68-6,7=11,3 | 127,69 | ||
| 52-56,7=-4,7 | 22,09 | ||
| 54-56,7=-2,7 | 7,29 | ||
| M=∑m:10=56,7 | ∑(m–M)=0 | ∑(m–M)2=434,1 |
2. У другу графу вписуємо варіанти (m).
3. Варіанти сумуємо, суму (∑) ділимо на число варіант і таким чином одержуємо середню арифметичну (М).
4. Віднімаємо М (середню арифметичну) від кожної m (варіанти) і різниці (відхилення від середньої) вписуємо у третю графу. Ці значення можуть бути від’ємними. Для перевірки правильності обчислень підсумовуємо всі різниці m–M, їхня сума повинна дорівнювати нулю.
5. Кожне відхилення підводимо до квадрату і вписуємо квадрати відхилень (m–M)2 у четверту графу. Квадрати відхилень завжди позитивні. Підсумовуючи всі числа четвертої графи, одержуємо суму квадратів відхилень ∑(m–M)2, яку вписуємо у підсумок четвертої графи.
6. За даними останнього рядка таблиці обчислюємо середньоквадратичне відхилення (s) за вище згаданою формулою: 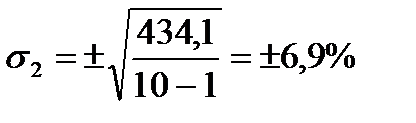
ПОХИБКА СЕРЕДНЬОЇ АРИФМЕТИЧНОЇ (m):
Знайдемо похибку середньої арифметичної (m) для партії меду №1:
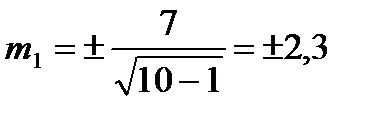
Знайдемо похибку середньої арифметичної (m) для партії меду №2:
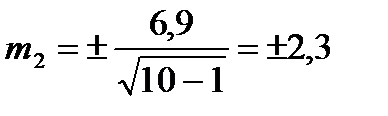
МІНІМАЛЬНЕ (Min) І МАКСИМАЛЬНЕ (Max) ЗНАЧЕННЯ:
Для партії меду №1 min=43%, max=67%.
Для партії меду №2 min=49%, max=68%.
КОЕФІЦІЄНТ ВАРІАЦІЇ (СV):
Знайдемо коефіцієнт варіації (С V) для партії меду №1: 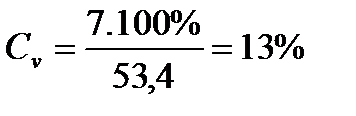
Знайдемо коефіцієнт варіації (С V) для партії меду №2: 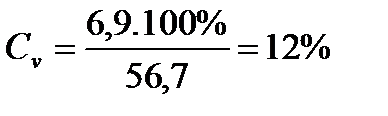
ОЦІНКА ДОСТОВІРНОСТІ РІЗНИЦІ МІЖ СЕРЕДНІМИ ВЕЛИЧИНАМИ ДВОХ ВИБІРОК:
Знайдемо достовірність різниці між середніми величинами вмісту пилкових зерен у меді партії №1 та партії №2:
1) Різниця (d) між двома середніми (M1 і M2): d = M1 – M2= 56,7-53,4=3,3
2) Середня помилка різниці (md): 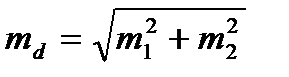
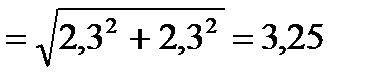
3) Вірогідність різниці (td): 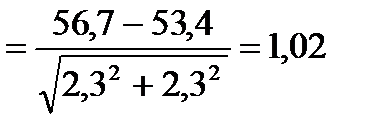
4) За таблицею 1 знаходимо достовірність Р: число ступенів свободи у нашій вибірці становить ν= (n1+ n2)–1 = (10+10)–1= 19. У рядочку, де стоїть число 19, знайдемо значення, яке було б найближче до знайденого нами td=1,02. Це число 1,73. Воно стоїть у стовпчику, де Р становить 0,1, а це означає, що різниця між двома вибірками не достовірна.
ПРИКЛАД РОЗВ’ЯЗУВАННЯ ТИПОВОГО ЗАВДАННЯ №2
Завдання 2. За нижче наведеними даними розрахувати коефіцієнт фенотипової кореляції між вмістом білка (х) і жиру (у) в молоці корів. Зробити відповідні висновки.
Вміст білка (х) і жиру (у) в молоці корів:
| х, % | 3,0 | 3,1 | 3,2 | 3,1 | 3,3 | 3,1 | 3,2 | 3,4 | 3,1 | 3,4 |
| у, % | 3,9 | 4,0 | 4,0 | 3,8 | 4,2 | 4,3 | 4,2 | 4,3 | 4,2 | 4,0 |
Розв’язання:
1) Складемо робочу таблицю та розрахуємо всі необхідні показники:
| Номер варіанти | х2 | х | у2 | у | (у-х)2 | (у-х) |
| 15,21 | 3,9 | 0,81 | 0,9 | |||
| 9,61 | 3,1 | 0,81 | 0,9 | |||
| 10,24 | 3,2 | 0,64 | 0,8 | |||
| 9,61 | 3,1 | 14,44 | 3,8 | 0,49 | 0,7 | |
| 10,89 | 3,3 | 17,64 | 4,2 | 0,81 | 0,9 | |
| 9,61 | 3,1 | 18,49 | 4,3 | 1,44 | 1,2 | |
| 10,24 | 3,2 | 17,64 | 4,2 | |||
| 11,56 | 3,4 | 18,49 | 4,3 | 0,81 | 0,9 | |
| 9,61 | 3,1 | 17,64 | 4,2 | 1,21 | 1,1 | |
| 11,56 | 3,4 | 0,36 | 0,6 | |||
| 101,93 | 1017,61 | 167,55 | 1672,81 | 8,38 | ||
| ∑х2 | (∑х)2 | ∑у2 | (∑у)2 | ∑d2 | (∑d)2 |
2) Розрахуємо Сх, Су, Сd:
 ;
; 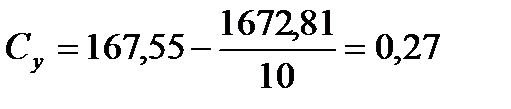 ;
; 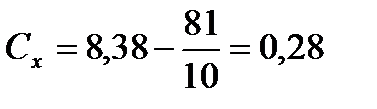
3) Розрахуємо коефіцієнт фенотипової кореляції за формулою:
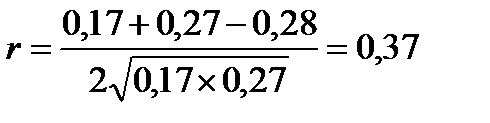
Відповідь: Коефіцієнт фенотипової кореляції становить +0,37, тобто міжвмістом білка і жиру в молоці корів існує малий позитивний зв'язок.
Date: 2015-09-02; view: 2239; Нарушение авторских прав