Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Сучасне уявлення про будову гена
|
|
У 1865 р. Г. Мендель на основі гібридологічного аналізу довів існування матеріальних носіїв спадковості, які він називав факторами, не розшифровуючи їх хімічного складу, функцій і місця знаходження в клітинах. Значно пізніше менделівські фактори за пропозицією В. Йогансена стали називати генами, але питання про їх хімічну природу і функції довго залишались відкритими. Після 1944 року, тобто після доведення ролі ДНК у спадковості, про ген стали говорити як про неподільну ділянку цієї молекули, в якій записана інформація про структуру конкретної білкової молекули. Таке визначення гена відображало тільки хімічну суть його будови. Але не дивлячись на це така уява про ген панувала довгий час.
Ген – це ділянка молекули нуклеїнової кислоти, яка характеризується специфічною послідовністю нуклеотидів і виконує функцію, яка відрізняється від функції інших генів та здатна змінюватися шляхом мутування (у хімічному відношенні, ген складається з нуклеотидів (у середньому 1-1,5 тис; ген тРНК – 190 нуклеотидів; ген фіброїну шовку тутового шовкопряда – 16 тисяч).
Якщо раніше вважали, що ген – це неподільна частина, то зараз доведено, що він складається з певних функціональних одиниць. Ген, як функціональну одиницю зараз називають цитроном, який поділяється на рекони і мутони.
Рекони – це ділянки гена, здатні до рекомбінації при кросинговері, мутони – здатні до мутації (мінливості). Розміри рекону та мутону досягають однієї або кількох пар, цистрону – сотні і тисячі нуклеотидів.
У геномі ссавців налічується 50-100 тисяч генів (геном – це сукупність генів у гаплоїдній клітині, тобто гаметі). Виявляється, що це значно більше, ніж може вмістити ДНК певного геному. Окрім того, навіть при такій кількості генів в організмі існує величезний генний дефіцит. Як відомо, на вторгнення кожного антигена організм відповідає синтезом специфічних антитіл проти них. Антитіла – це білки, так звані імуноглобіни, кожний з яких повинен кодуватись своїм геном.
Виходить, що тільки генів, які повинні працювати на імунну систему, повинно бути стільки, скільки могло б існувати антигенів – безліч та плюс гени, які відповідають за видові ознаки. У чому полягає секрет такої невідповідності: у неправильному підрахунку кількості генів чи в чомусь іншому?
Дослідженнями останніх років було встановлено, що у геномі є два типи генів:
ü постійні, або стабільні (приблизно половина генів) – гени, які мають чітко визначену послідовність нуклеотидів (може змінюватись тільки у результаті мутацій), що визначають видові ознаки організму (наприклад, зовнішній вигляд);
ü непостійні, або лабільні – гени, послідовність нуклеотидного складу яких, у міру потреби організму, може змінюватись за рахунок тасування екзотів та нітронів, з яких вони складаються.
| Екзонипредставлені ланцюгом нуклеотидів, в яких закодована відповідна генетична інформація. Інтронискладаються з нуклеотидів, що не несуть генетичної інформації (схематично: … – екзон – інтрон – екзон – нітрон – …). Довжина інтронних ділянок варіює від 10 до 20 тис. пар нуклеотидів, що за сумарною довжиною нерідко в кілька разів перевищує довжину екзонних ділянок. Між екзонома та інтронами може відбуватися тасування ділянок. Кожна така перестановка гена призводить до зміни його функції. Це означає, що один і той самий ген, маючи кілька варіантів перебудови, дає нову функцію, тобто новий імуноглобулін-антитіло. Якщо можливості в перебудові кожного гена помножити на кількість генів, то варіантів буде дуже багато. Цим і пояснюється «парадокс» з кількістю генів у геномі і їх можливостями. Особливості зчитування інформації з лабільних генів (етапи синтезу): ü транскрипція на ДНК (гені) молекули-попередника, або про-іРНК; ü процесинг, у ході якого за участі ферментів рестриктаз вирізуються інтронні ділянки; ü сплайсинг, у ході якого за участі ферментів лігаз зшиваються екзонні ділянки й утворюються «зрілі» молекули іРНК, які виходять з ядра і потрапляють у рибосоми, де і відбувається синтез білка. | 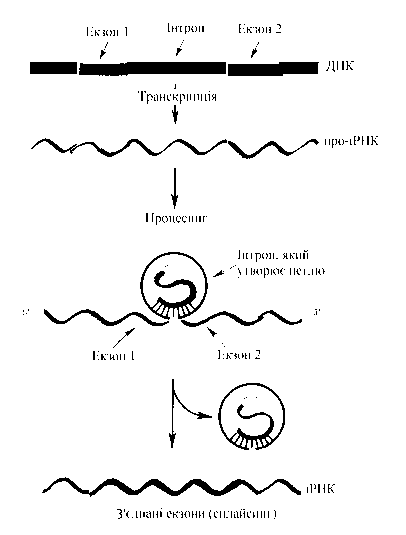
|
Механізм пристосувального характеру еволюції
У структурі генів закладена важлива риса еволюції – її доцільність. До відкриття екзонно-інтронної будови генів неможливо було відповісти на основне питання еволюції: в чому полягає механізм пристосувального характеру еволюції і які механізми її швидкого поступального руху? Сьогодні це пояснюється дуже просто. Умови зовнішнього середовища, які постійно змінюються, змінюють і внутрішнє середовище (склад цитоплазми і т. д.), яке впливає на перебудовні процеси в лабільних генах. Зрозуміло, що відповідно до середовищних запитів гени, перебудовуючись, видають потрібну інформацію і змінюють функції окремих процесів, органів тощо. Крім таких функціональних змін генів, існували й існують мутації, які на тривалий час закріплюють частоповторювані зміни в структурі генів.
Є відомості про те, що гени можуть перебувати як у межах однієї клітини, так і поза межами клітин, тканин і навіть організму. Прикладом таких генів є онкогени, що можуть перетворюватись на онковіруси. Крім того, відомо, що є так звані «транспозони», здатні змінювати своє положення в хромосомі. Транспозиція – це переміщення з місця на місце в хромосомі невеликих фрагментів ДНК (генів). Вважають, що вони відіграють важливу роль як в еволюції генетичного матеріалу, так і в регуляції дії генів, у тому числі диференціації клітин і тканин організму, що розвивається. Механізми тасування всередині генів, утворення мільйонів нових генів, транспозиції, створені природою, і визначають індивідуальність організму.
Отже, ген – це не просто ділянка ДНК, а складна система, яка має механізми до перебудови, в результаті чого змінюються його функції. Хоч багато вже відомо про ген, але його вивчення ніколи не припиниться, тому що він невичерпний, як і саме життя.
4. Генетичний код та його властивості
Генетичний код – принцип запису генетичної інформації на гені (повністю розшифрований).
У 1964 р. Дж. Маттей, Ф. Крік, С. Очоа і М. Ніренберг розшифрували генетичний код і встановили його властивості.
Властивості генетичного коду:
Триплетність: генетична інформація в гені записана триплетами. Триплет (кодон, кодоген) – три сусідніх нуклеотиди в нуклеїнових кислотах, що несуть інформацію на включення однієї з 20 основних амінокислот у поліпептидний ланцюг молекули білка. Ініціюючі кодони чи стоп-кодони (безглузді) − триплети, які кодують початок чи кінець синтезу білка.
Виродженість: кожній амінокислоті відповідає не один кодон, а кілька (наприклад, амінокислоту фенілаланін кодують два триплети: УУУ, УУЦ, треонін – чотири: АЦУ, АЦЦ, АЦА, АЦГ).
Квазідуплетність: основну генетичну інформацію у коді несе дуплет, що стоїть на його початку (наприклад, треонін − АЦ У, АЦ Ц, АЦ А, АЦ Г).
Колінеарність: амінокислоти розміщені в білковій молекулі у такій послідовності, в якій кодони, що їх кодують, знаходяться у гені.
Лінійність (неперекриваючість): зчитування спадкової інформації з ДНК на іРНК відбувається в одному напрямку триплет за триплетом.
Універсальність: генетичний код однаковий для всіх живих організмів (вірусів, бактерій, грибів, рослин, тварин).
*Повний словник генетичного коду для амінокислот (наведено триплети іРНК)
| Перша літера | Друга літера | Третя літера | ||||||||
| У | Ц | А | Г | |||||||
| У | УУУ УУЦ | (фен) фенілаланін | УЦУ УЦЦ УЦА УЦГ | (сер) серин | УАУ УАЦ | (тир) тирозин | УГУ УГЦ | (цис) цистин | У Ц А | |
| УУА УУГ | (лей) лейцин | УАА УАГ | (стоп-кодон) кінець синтезу | УГА | (стоп-кодон) кінець синтезу | |||||
| УГГ | (три) триптофан | Г | ||||||||
| Ц | ЦУУ ЦУЦ ЦУА ЦУГ | (лей) лейцин | ЦЦУ ЦЦЦ ЦЦА ЦЦГ | (про) пролін | ЦАУ ЦАЦ | (гіс) гістидин | ЦГУ ЦГЦ ЦГА ЦГГ | (арг) аргінін | У | |
| Ц | ||||||||||
| А | ||||||||||
| Г | ||||||||||
| ЦАА ЦАГ | (глун) глутамін | |||||||||
| А | АУУ АУЦ АУА | (ілей) ізолейцин | АЦУ АЦЦ АЦА АЦГ | (тре) треонін | ААУ ААЦ | (аспн) аспарагін | АГУ АГЦ | (сер) серин | У | |
| Ц | ||||||||||
| ААА ААГ | (ліз) лізин | АГА АГГ | (арг) аргінін | А | ||||||
| АУГ | (мет) метіонін (ініціюючий кодон) | Г | ||||||||
| Г | ГУУ ГУЦ ГУА ГУГ | (вал) валін | ГЦУ ГЦЦ ГЦА ГЦГ | (ала) аланін | ГАУ ГАЦ | (асп) аспарагінова к-та | ГГУ ГГЦ ГГА ГГГ | (глі) гліцин | У | |
| Ц | ||||||||||
| ГАА ГАГ | (глу) глютамінова к-та | А | ||||||||
| Г |
*За допомогою цієї таблиці можна визначити, яку саме амінокислоту кодує певний триплет. Перший нуклеотид у триплеті беруть із лівого вертикального стовпчика, другий − з верхнього горизонтального і третій − з правого вертикального. В місці перетину ліній знаходиться інформація про амінокислоту, яку слід визначити.
5. Загальна характеристика білків та їх біосинтез
5.1. Загальна характеристика білків
Білки– високомолекулярні біополімери, мономерами яких є амінокислоти.
Мономери (20 амінокислот) мають загальну формулу (всі амінокислоти подібні карбоксильною (–СООН – кислотні властивості) та аміногрупою (–NH2 – лужні властивості), відмінні – бічними ланцюгами – радикалами (або R-групами).
| 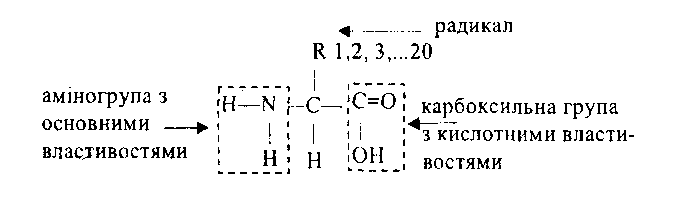 Схема будови амінокислоти
Схема будови амінокислоти
|
Амінокислоти поділяються на:
незамінні* – не можуть синтезуватись тваринними організмами з інших сполук і тому обов'язково мають надходити ззовні (з їжею) (лізин, триптофан, фенілааанін, метіонін, треонін, лейцин, ізолейцин, валін).
замінні – синтезуються у тваринних організмах у достатній кількості з незамінних амінокислот або інших сполук.
Білки класифікують:
1) залежно від продуктів розпаду:
ü прості (протеїни) – складаються лише з амінокислот (альбуміни, глобуліни – білки крові; склеропротеїни – кератин волосся, пір’я; колаген – сухожилля);
ü складні (протеїди) – мають у складі не лише амінокислоти, а і небілковий матеріал – простетичну групу (нуклеопротеїди (сполучення нуклеїнових кислот і простих білків) – хромосоми; хромопротеїди (забарвлені речовини небілкової природи і прості білки) гемоглобін, цитохром; фосфопротеїди – казеїн молока; глікопротеїди – плазма крові, муцин слини тощо).
2) залежно від вмісту амінокислот: повноцінні – містять усі незамінні амінокислоти; неповноцінні – не містять тих чи інших незамінних амінокислот.
Амінокислоти, які входять у склад білків і їх умовні позначення
| Назва амінокислоти | |||
| Повна | Скорочена | Повна | Скорочена |
| Аланін | ала | Лейцин* | лей |
| Аргінін (незамінна для дітей) | арг | Лізин* | ліз |
| Аспарагін | аспн | Метіонін* | мет |
| Аспарагінова кислота | асп | Пролін | про |
| Валін* | вал | Серин | сер |
| Гістидин (незамінна для дітей) | гіс | Тирозин | тир |
| Гліцин | глі | Треонін* | тре |
| Глутамін | глун | Триптофан* | три |
| Глутамінова кислота | глу | Фенілаланін* | фен |
| Ізолейцин* | ілей | Цистеїн | цис |
5.2. Біосинтез білка є результатом реалізації генетичної інформації у клітині, яка здійснюється за схемою:
Ген (ДНК)
іРНК
Білок
Ознака
транскрипція
трансляція
Етапи біосинтезу білка
| Етап | Процес | Де відбувається | ||||||||||
Транскрипція (від лат. транскриптіо − переписування) – процес переписування інформації з ДНК на іРНК. Під час цього процесу відбувається синтез іРНК (мРНК) на ДНК за участю ферменту ДНК-залеженої-РНК-полімерази за принципом комплементарності:
Спочатку синтезується про-іРНК, яка знімає копію з усіх екзонних та інтронних ділянок ДНК, після чого за допомогою спеціальних ферментів з неї видаляються ділянки, які не несуть генетичної інформації (інторони) і вона перетворюється на активну форму. | У ядрі на ДНК. (після синтезу іРНК мігрує до рибосом у цитоплазму). | |||||||||||
| Трансляція (віл лат. транслятіо − передача) − перенесення інформації, записаної у послідовності нуклеотидів іРНК, у певну послідовність амінокислот називається (синтез білка на іРНК). У цьому процесі виділяють три стадії: Ø активація амінокислот; Ø аміноацелювання тРНК; Ø власне трансляція. | У цитоплазмі на рибосомах | |||||||||||
| 2.1. Активація амінокислот Амінокислоти під впливом ферменту (аміноацил-тРНК-синтетаза), специфічного для кожної амінокислоти взаємодіють з АТФ у результаті чого утворюються аміноациладенилати (комплекс ферменту аміноацил-тРНК-синтетази та амінокислотного залишку). 2.2. Аміноацелювання тРНК Аміноациладенилати приєднуються до молекули специфічної тРНК і мігрують до рибосом. Певна амінокислота переносить певною тРНК, що визначається антикодоном. Антикодон − триплет на середній лопаті (верхівці) специфічної тРНК, комплементарний відповідному триплету (кодону) іРНК. | У цитоплазмі. | |||||||||||
| 2.3. Власне трансляція Аміноациладенилати, згідно словника генетичного коду, полімеризуються у поліпептидний ланцюг. У цьому процесі виділяють три стадії: Ø ініціацію – початок синтезу поліпептидного ланцюга (приєднання першої амінокислоти); Ø елонгацію – подовження поліпептидного ланцюга (приєднання амінокислот); Ø термінацію – закінчення синтезу і відщеплення готового поліпептиду (приєднання останньої амінокислоти). | На рибосомах (шорсткій або гранулярній ЕПС). Синтезована молекула білка надходить у порожнину ендоплазматичної сітки, якою транспортується в певну ділянку клітини. | |||||||||||
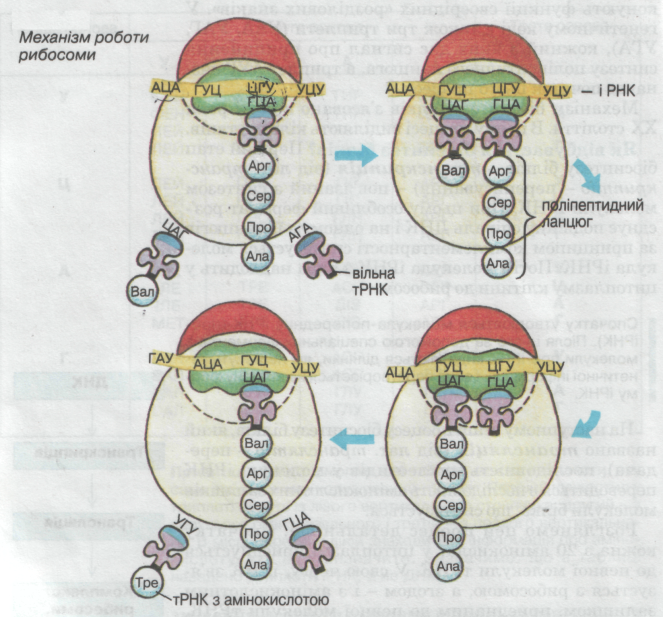
| Схема трансляції (механізм роботи рибосоми): 1) Рибосома насувається на ниткоподібну молекулу іРНК таким чином, що іРНК опиняється між двома її субодиницями. 2) Рибосома «ковзає» зліва направо по молекулі іРНК і збирає молекулу білка. Кожен крок рибосоми дорівнює одному триплету. 3) Коли рибосома дещо просунеться вперед по молекулі іРНК, на її місце надходить друга, а згодом − третя, четверта тощо і біосинтез нових білкових молекул триває далі. Кількість рибосом, які одночасно можуть бути розташовані на молекулі іРНК, зумовлена довжиною останньої. 4) Коли рибосома досягає одного з трьох триплетів (УАА, УАГ, УГА), що сигналізує про припинення синтезу поліпептидного ланцюга, вона разом із білковою молекулою залишає іРНК. 5) Рибосома розпадається на субодиниці, які потрапляють на будь-яку іншу молекулу іРНК. Процеси біосинтезу білка належать до реакцій матричного синтезу, тому що з ДНК, потім з іРНК знімається інформація як з матриці. | |||||||||||
| 2.4. Дозрівання білка Синтезований білок набуває своєї природної просторової (вторинної, третинної) структури і функціональної активності: за участю відповідних ферментів від нього відщеплюються зайві амінокислотні залишки, вводяться небілкові фосфатні, карбоксильні та інші групи, приєднуються вуглеводи, ліпіди тощо. | У цитоплазмі | |||||||||||
6. Регуляція активності генів
Залежно від фази розвитку клітини, умов існування та інших факторів одночасно працюють лише 5 % генів, а 95 % – знаходяться в неактивному стані. Крім того, ці 5 % активних генів не одні і ті самі.
Черговість включення і виключення генів регулюється в організмі. Складна регуляція активності генів забезпечує найефективніше використання ресурсів організму, дає змогу уникнути затрат енергії і пластичних матеріалів на синтез сполук, непотрібних у даний час. З економічної точки зору надлишок синтезу речовин може бути не тільки невигідний, а навіть шкідливий для організму. Отже, успадковується не тільки геном (повний набір генів), а й система, що регулює його діяльність.
У 1961 році французькі вчені Ф. Жакоб і Ж. Моно запропонували схему регуляції активності генів (на прикладі кишкової палички).
Узагальнивши велику кількість даних, вони зробили висновок, що в генній системі кишкової палички є три види генів:
Ø структурні – визначають послідовність амінокислот у поліпептидному ланцюгу;
Ø функціональні (гени-оператори) – контролюють функцію структурних генів;
Ø гени-регулятори – регулюють діяльність генів-операторів.
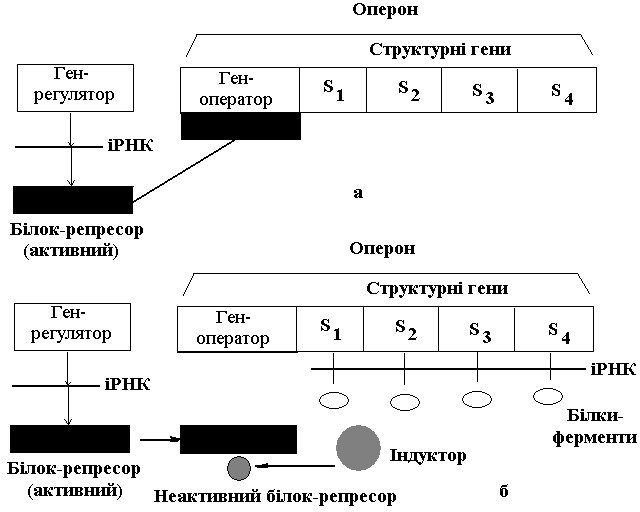
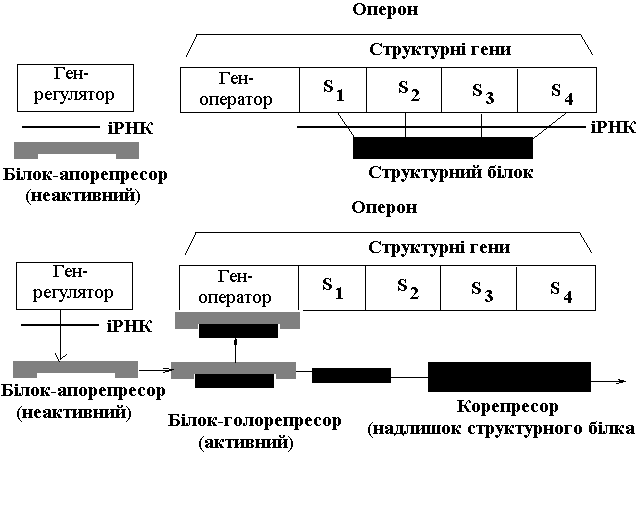
Ген-оператор, ряд структурних генів (S1, S2, S3, S4), розміщених поряд у лінійній послідовності становлять так званий оперон. Система генів, у яку входять декілька (4-5) структурних генів, ген-оператор і ген-регулятор здійснюють регуляцію синтезу конкретного одного білка-ферменту.
В організмі виробляються пластичні білки і білки-ферменти. Пластичні білки призначені для побудови тіла, білки-ферменти є каталізаторами біохімічних реакцій в організмі. Тому регуляція синтезу білків-ферментів і пластичних білків здійснюється за двома різними схемами.
Регуляція синтезу білків-ферментів
В основі регуляції синтезу білків-ферментів лежить два взаємопротилежних процеси – індукція (збудження синтезу) та репресія (пригнічення синтезу). Як індуктори (збуджувачі) виступають поживні речовини (жири, білки, вуглеводи тощо), що надходять в організм з кормом, для розщеплення яких потрібні відповідні ферменти. Репресорами (пригнічувачами) є активні білки-репресори, які кодуються генами-регуляторами.
| Регуляції активності генів під час синтезу білків-ферментів (за Ф.Жакобом і Ж.Моно) | Регуляції активності генів під час синтезу пластичних білків (за Ф.Жакобом і Ж.Моно) |
| Ген-регулятор постійно видає інформацію на синтез активного білка-репресора, який, зв'язуючись з геном-оператором, блокує його і заважає включенню структурних генів, що не видають ніякої інформації. Коли в клітину поступає індуктор (поживна речовина, для розщеплення якої потрібні певні ферменти), він зв'язує активний білок-репресор та інактивує його. Ген-оператор, не зв’язавшись з білком-репресором, стає активним і включає структурні гени. Структурні гени видають інформацію на синтез білків-ферментів, які розщеплюють поживні речовини. Як тільки індуктор повністю розщепиться, білок-репресор стає активним і блокує ген-оператор, внаслідок чого відключаються структурні гени. Отже, гени, які відповідають за синтез білків-ферментів включаються і виключаються залежно від надходження і розщеплення індуктора. | Ген-регулятор постійно видає інформацію на синтез білка-апорепресора (неактивного білка), який не може з'єднуватись з геном-оператором. Тому ген-оператор включає структурні гени, які видають інформацію на синтез пластичного білка доти, поки його не буде в надлишку. При перевиробництві пластичного білка його надлишок виступає як корепресор, який сполучається з білком-апорепресором і перетворює його в активний голорепресор, який блокує ген-оператор, а той відключає структурні гени. Як тільки надлишок пластичного білка клітина використає, білок-апорепресор стає неактивним і система генів знову працює на синтез пластичного білка. Тобто, гени, які відповідають за синтез пластичних білків включаються і виключаються залежно від потреб клітини у пластичних білках. |
| 
|
Отже, успадковується не тільки геном, а й система, що регулює його діяльність. Вона забезпечує почергове включення певних генів, тому найефективніше використання ресурсів організму.
МОДУЛЬ 2. СПЕЦІАЛЬНА ГЕНЕТИКА
Лекція 8. МІНЛИВІСТЬ ОРГАНІЗМІВ ТА БІОМЕТРИЧНІ МЕТОДИ ЇЇ ВИВЧЕННЯ
План:
1. Мінливість організмів та її види.
2. Закон гомологічних рядів спадкової мінливості М.І.Вавилова та його практичне значення для селекції.
3. Біометричні методи вивчення мінливості.
1. Мінливість організмів та її види.
Мінливість – це здатність живих істот, їх ознак і властивостей змінюватись у визначених межах, у результаті чого виникають деякі відмінності між особинами і групами особин, навіть родинноблизькими.
| Види мінливості |
| Неспадкова (фенотипова, визначена) |
| Спадкова (генотипова, невизначена) |
| Модифікаційна |
| Мутаційна Комбінаційна Кореляційна |
Порівняльна характеристика видів мінливості
| Вид мінливості | Неспадкова (фенотипова, визначена) | Спадкова (генотипова, невизначена) | ||
| Модифікаційна | Мутаційна | Комбінаційна | Кореляційна | |
| Визначення | Сукупність змінених ознак і властивостей певної особини, спричинені змінами умов середовища життя і не пов’язані зі змінами генотипу. | Сукупність змінених ознак і властивостей певної особини, які зумовлені мутаціями. | Сукупність змінених ознак і властивостей певної особини, які зумовлені утворенням у її генотипі нових сполучень генів через комбінаторику спадкової інформації. | Сукупність змінених ознак і властивостей певної особини, які зумовлені зміною інших ознак, зв’язаних з нею фізіологічно та анатомічно. Розрізняють кореляцію: позитивну – посилення розвитку однієї ознаки призводить до посиленого розвитку іншої; негативну – посилення розвитку однієї ознаки гальмує розвиток іншої. |
| Причини виникнення | Вплив умов середовища на прояв генотипу. | Мутації – стійкі зміни генотипу, які виникають раптово під впливом мутагенів і призводять до зміни спадкових ознак організму. | ü кросинговер гомологічних хромосом у профазі І мейозу; ü незалежне розходження хромосом в анафазі І мейозу; ü випадкове поєднання алельних генів при злитті гамет. | Вплив умов середовища на розвиток певної ознаки, яка тягне за собою зміну іншої, зв’язаної з нею фізіологічно та анатомічно. |
| Приклад | 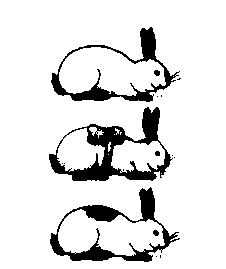  Розвиток горностаєвого забарвлення шерсті кролів залежно від температури навколишнього середовища.
Розвиток горностаєвого забарвлення шерсті кролів залежно від температури навколишнього середовища.
| Відсутність вушної раковими у овець.

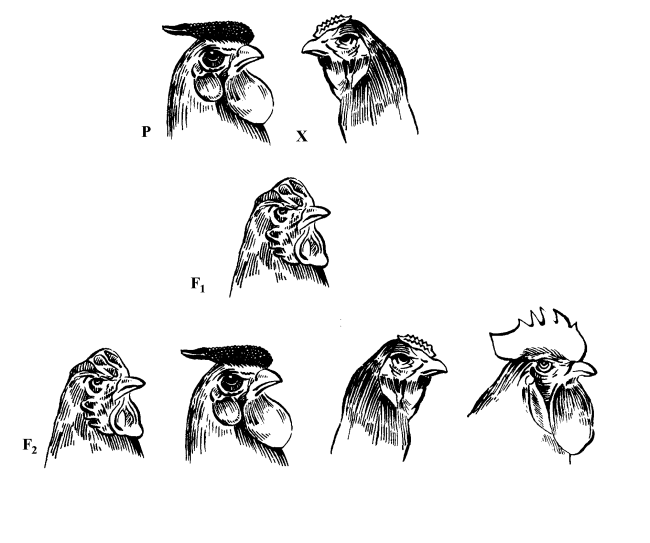
| Формування різних форм гребеня у курей залежно від наявності генів у генотипі.
| Позитивна кореляція (чим більше вим’я у корів, тим більший надій молока). Негативна кореляція (при збільшенні надою у корів зменшується жирність молока) |
Неспадкова (модифікаційна) (від лат. модус – міра, вигляд і фаціо – роблю) мінливість – зміни ознак організму (його фенотипу), спричинені змінами умов середовища життя і не пов’язані зі змінами генотипу.
Модифікаційні зміни (модифікації) – це реакції організмів на зміну інтенсивності дії певних чинників довкілля. Вони однакові для всіх генотипно однорідних організмів.
Причини виникнення: Зумовлена впливом середовища на прояв генотипу.
Конкретний алельний ген зумовлює не розвиток певного, кодованого ним стану ознаки, а лише межі (норму реакції), в яких той може змінюватися залежно від інтенсивності дії тих чи інших чинників навколишнього середовища.
Норма реакції – це межі модифікаційної мінливості ознак, зумовлені генотипом організму (або, будь-яка ознака може змінюватись лише в певних межах).
Властивості модифікацій:
· ступінь вираження модифікацій прямо залежить від інтенсивності й тривалості дії на організм певного чинника (у рачка-артемії ступінь волохатості задньої частини черевця залежить від солоності води: тим більша, чим нижча концентрація солей у воді);
· модифікації не успадковуються (довів німецький учений А. Вейсман, який протягом багатьох поколінь відрізав мишам хвости, і з’ясував, що у безхвостих батьків завжди народжувалися хвостаті нащадки);
· модифікації можуть зникати протягом життя однієї особини, якщо припиняється дія чинника, який їх викликав (загар, набутий людиною влітку, поступово зникає протягом осінньо-зимового періоду);
· деякі модифікації, які виникли переважно на ранніх етапах індивідуального розвитку, можуть зберігатися протягом усього життя особини (викривлення кісток нижніх кінцівок унаслідок рахіту зберігається протягом усього життя).
· модифікації виникли в процесі історичного розвитку виду як певні пристосувальні реакції на зміни умов середовища існування, з якими постійно стикаються організми;
· деякі модифікації не мають пристосувального характеру (якщо затінити нижню частину стебла картоплі, то на ній почнуть утворюватися надземні бульби), вони виникають тоді, коли організми опиняються в незвичних для себе умовах, з якими не доводилося постійно стикатися їхнім предкам.
Типи ознак залежно від межі норми реакції:
· ознаки, які визначають життєздатність організмів (взаєморозташування внутрішніх органів) – мають найвужчі межі норми реакції;
· ознаки, які не визначають життєздатність організмів (маса тіла, ріст, колір волосся) – мають широкі межі норми реакції;
Типи ознак залежно від впливу на них умов середовища:
· ознаки, стани яких майже повністю визначаються генотипом (розташування очей, кількість пальців на кінцівках, група крові, характер жилкування листків тощо)
· ознаки, стани яких дуже визначаються умовами довкілля (ріст і маса організмів, розміри листкової пластинки, розвиток горностаєвого забарвлення шерсті кролів залежить від температури тощо)
Порівняння модифікаційної і мутаційної мінливості
| Модифікаційна | Мутаційна |
| Пов’язані із впливом середовища | |
| Нащадками не успадковується | Нащадками успадковується |
| Пов’язана з фенотипом і на генотип не впливає. | Пов’язані з перебудовою спадкового матеріалу (генів, хромосом, кількості хромосом). |
| З’являються масово. Має варіаційний ряд і середню величину. | З’являються лише в окремих особин. |
| Має спрямований характер. | Мають неспрямований характер (однакові мутації можуть виникати у результаті впливу різних факторів, і навпаки, одні фактори можуть викликати різні мутації). |
| Мають пристосувальний характер. | Пристосувального характеру не мають (можуть бути шкідливими, нейтральними, шкідливими). |
| Сприяє пристосуванню виду до мінливих умов середовища і його збереженню. | Дає матеріал для природного добору і еволюції (нейтральні мутації за певних змін середовища існування можуть виявитися корисними для організмів). |
| Знання закономірностей модифікаційної мінливості у селекції дозволяє підбирати такі умови, за яких у фенотипі організмів проявляються корисні для людини стани ознак і пригнічуються прояви шкідливих. | Використання мутацій у селекції рослин і мікроорганізмів збільшує різноманітність вихідного матеріалу і підвищує ефективність селекційної роботи. |
Практичне значення мінливості у тваринництві:
ü створюють нові породи тварин;
ü шляхом підбору батьківських пар здійснюється виправлення у нащадків недоліків одного з батьків позитивними якостями іншого.
Велике значення мінливості у штучному доборі відмічали такі вчені як М.Ф. Іванов, П.Н. Кулешов, І.В Мічурін, М.І.Вавилов.
2. Закон гомологічних рядів спадкової мінливості М.І.Вавилова та його практичне значення для селекції (1920 р.)
| Генетично близькі види і роди характеризуються подібними рядами спадкової мінливості з такою правильністю, що, вивчивши ряд форм у межах одного виду або роду, можна передбачити наявність форм із подібним поєднанням ознаку межах близьких видів або родів. При цьому, чим тісніші родинні зв'язки між організмами, тим подібніші ряди їхньої спадкової мінливості. Цю закономірність М.І. Вавилов встановив для рослин, але вона виявилась універсальною для всіх організмів. Генетична основа закону: ступінь історичної спорідненості організмів прямо пропорційний кількості їхніх спільних генів, тому і мутації цих генів можуть бути подібними. У фенотипі це проявляється подібною мінливістю багатьох ознак у близьких видів, родів та інших таксонів. |  Гомологічні ряди мінливості за остюками: 1-4 − м'яка пшениця; 5-8 − тверда пшениця; 9-12 − шестирядний ячмінь. Форми з довгими (1, 5, 9) і короткими (2, 6, 10) остюками, здуті (3, 7, 11) і без остюків (4, 8, 12).
Гомологічні ряди мінливості за остюками: 1-4 − м'яка пшениця; 5-8 − тверда пшениця; 9-12 − шестирядний ячмінь. Форми з довгими (1, 5, 9) і короткими (2, 6, 10) остюками, здуті (3, 7, 11) і без остюків (4, 8, 12).
|
Практичне значення закону:
- дає змогу передбачити можливість появи мутацій, ще не відомих науці, які можна використовувати у селекції для створення нових цінних для народного господарства сортів рослин, порід тварин та штамів мікроорганізмів;
- пояснює спрямованість історичного розвитку споріднених груп організмів;
- у систематиці організмів дає можливість передбачити існування невідомих науці систематичних груп (видів, родів тощо) з певною сукупністю ознак, якщо такі форми виявлено в споріднених групах.
Деякі спадкові дефекти людини та тварин
| Спадкові аномалії | Людина | Велика рогата худоба | Свині | Вівці | Коні | Собаки | Кури | Гризуни |
| Артрогрипоз (анкілоз або контрактура м’язів) | + | + | + | + | + | + | + | + |
| Карликовість | + | + | + | + | + | + | + | + |
| Відсутність кінцівок | + | + | + | + | + | + | ||
| Атаксія (порушення координації рухів) | + | + | + | + | + | + | + | + |
| Полідактилія | + | + | + | + | + | + | ||
| Іхтіоз | + | + | + | + | + | |||
| Гемофілія | + | + | + | + | + |
Наприклад, альбінізм зареєстрований у багатьох видів (людини, овець, кіз, собак, кішок, кролів, мишей і ін.). Ця аномалія повинна бути виявлена і у тих видів, у яких до теперішнього часу невідома. Генетичний паралелізм у відношенні спадкових аномалій виявлений у людини і багатьох видів тварин. У людини відомо більше 2500 спадкових аномалій. У тварин генетичних аномалій відомо на багато менше. Виходячи із закону гомологічних рядів, у майбутньому подібні аномалії будуть виявлені і у сільськогосподарських тварин.
3. Біометричні методи вивчення мінливості.
Мінливість, яка спостерігається як серед диких, так і серед домашніх тварин і рослин, вимагає спеціальних методів її вивчення, необхідних для характеристики цілої групи особин, тому що мінливість окремих індивідів не може охарактеризувати цілу групу.
Розробленням методів вивчення мінливих ознак займається спеціальна наука – біометрія (bios – життя, metreo – вимірюю), математичною основою якої є теорія імовірності.
3.1. Поняття про якісні і кількісні ознаки
Тварини мають величезну різноманітність морфологічних, фізіологічних та господарсько-корисних ознак. Багато з них мають значення для практики тваринництва, на покращання і удосконалення яких спрямована племінна робота. Всі ознаки поділяють на якісні і кількісні.
До якісних ознак тварин відносяться: стать, колір хутрового покриву, тип ВНД, тілобудови і т.ін. Багато якісних ознак мають тільки два можливих альтернативних стани, наприклад стать жіноча чи чоловіча, а деякі – 3-4 (тип тілобудови, ВНД і т.ін.). Окремі якісні показники можуть мати кількісне вираження. Наприклад, ступінь пігментації хутра, ступінь вгодованості тварин можна оцінити балами (1, 2, 3 і т.д.). Однак для характеристики показників, як правило, достатньо провести окомірну оцінку і дати словесний опис їх у конкретної тварини або у групи тварин.
Кількісні ознаки відрізняються тим, що можуть бути виміряні і виражені у кілограмах, сантиметрах, відсотках і т. ін. До кількісних показників відносяться: надій, вміст жиру і білка у молоці, жива маса тварин, вік, плодючість і т.ін. Перехід від одного кількісного показника до другого становить безперервний ряд величин.
3.2. Генеральна і вибіркова сукупності
Біометрія дозволяє вивчати варіюючу ознаку на масовому матеріалі, наприклад на всіх тваринах даної лінії, породи, стада. Такий масовий матеріал називається генеральною сукупністю. Наприклад, генеральною сукупністю може бути вся чисельність тварин певного виду у нашій країні. Цю сукупність вивчають шляхом проведення державного перепису з урахуванням показників, які цікавлять (порода, вік, стать, продуктивність). Одержані у результаті перепису дані опрацьовують і визначають різні біометричні характеристики.
Однак вивчення генеральних сукупностей при великій чисельності тварин – складний і дорогоцінний захід. Тому застосовують метод вибікового дослідження, який дозволяє оцінити генеральну сукупність шляхом відбору меншої чисельності досліджених тварин.
Вибіркова сукупність повинна правильно відображати якості і особливості тварин, які становлять генеральну сукупність. Така умова забезпечується відбором частини тварин із генеральної сукупності за принципом випадкової вибірки.
Для ряду ознак вивчення їх варіювання у всіх особин генеральної сукупності неможливе ще і тому, що це може призвести до її знищення. У таких випадках вибіркова сукупність – єдиний спосіб, який дозволяє вивчити ту чи іншу ознаку.
Прикладом вибіркових проб є середні проби молока, крові, зерна, меду, м’яса і т.ін.
Отже, предметом біометрії є варіююча ознака, а основним її методом служить випадкова вибірка об’єктів для детального математичного (біометричного) аналізу.
3.3. Варіаційний ряд та варіаційна крива.
Варіаційний (від лат. варіатіо – зміна) ряд – послідовність кількісних показників проявів станів певної ознаки (варіант), розташованих у порядку їхнього зростання чи зменшення.
Аналіз варіаційного ряду:
· довжина варіаційного ряду свідчить про розмах модифікаційної мінливості;
· чим коротший варіаційний ряд, тим стабільніші умови навколишнього середовища, і навпаки;
· найбільша кількість варіант розташована в середній його частині, тобто вони мають середнє значення певної ознаки, тому що мінімальні та максимальні значення розвитку певної ознаки формуються тоді, коли більшість чинників довкілля діє в одному напрямі: найбільш або найменш сприятливому;
· ступінь розвитку певної ознаки у більшості особин виду усереднені, тому що одні фактори сприяють розвитку ознаки, інші, навпаки, гальмують (більшість людей має середній зріст, і лише незначна частина серед них – велетні або карлики).

| 
|
| Варіаційний ряд листків лавровишні | Варіаційна крива кількості колосків у колосі пшениці |
Варіаційна крива – це графічне зображення кількісних показників мінливості певної ознаки, яке ілюструє межі модифікаційної мінливості та частоту зустрічальності окремих варіант. За допомогою варіаційної кривої можна встановити середні показники і норму реакції певної ознаки.
3.4. Середні величини
Основними статистичними параметрами, які характеризують середній рівень варіюючої ознаки в генеральній чи вибірковій сукупності, служать величини середніх значень ознаки, які позначаються буквами латинського алфавіту: середня арифметична (X, M), середня геометрична (G), середня квадратична (S), середня гармонічна (Н), мода (Mo) та медіана (Me).
Перераховані параметри є показниками середнього рівня ознаки, яка варіює у межах від мінімального (xmin) до максимального (xmax) значення.
Залежно від поставленого завдання застосовують той чи інший параметр. Для характеристики кількісних ознак найчастіше використовують середню арифметичну.
Якщо потрібно визначити ознаку, яка характеризує величину площі круга або об’єм кулі (діаметр або площа еритроцитів, об’єм жирової кульки, об’єм клітинного ядра), то використовують середню квадратичну.
У випадку визначення середнього приросту живої маси тварин за періодами онтогенезу обчислюють середню геометричну (G).
При встановленні середньої швидкості бігу коня, швидкості молоковіддачі (тобто коли збільшення ознаки виражають оберненою величиною витраченого часу) обраховують середню гармонічну (Н).
Медіана (Ме) та мода (Mo) є допоміжними характеристиками середнього значення варіюючої ознаки у сукупності. Мода показує, яка величина варіанти даної ознаки найчастіше зустрічається у сукупності. Медіана вказує на те, який варіант розміщений у середині (центрі) варіаційного ряду, цей варіант ділить сукупність на дві рівні частини.
Співвідношення між усіма розглянутими середніми параметрами закономірне і виражається таким чином:
Mo
xmin< Н < G< X< S< xmax
Me
Тобто, величина середньої квадратичної (S) завжди більша, а величини середньої гармонічної (Н) та середньої геометричної (G) завжди менші будь-якої іншої середньої. У нормальному розподілі Х, Me та Mo співпадають. Неправильно вибраний параметр спотворює істину середню величину ознаки.
3.5. Показники мінливості (варіювання) ознаки
При вивченні варіювання ознаки особин певної сукупності застосовують такі основні статистичні параметри, як ліміт (ліміт= xmax– xmin), середньоквадратичне відхилення (s), коефіцієнт варіації (Cv %), варіансу (s2), нормальне відхилення (t).
Найпростіший показник варіювання ознаки – ліміт, тобто абсолютна різниця між максимальною і мінімальною величиною ознаки. Чим більша величина ліміту, тим значніше варіювання ознаки.
Основним критерієм мінливості є середньоквадратичне відхилення (s), яке показує, наскільки в середньому відхиляється кожен член сукупності від середньої арифметичної певного показника даної сукупності. Величина сигми завжди іменована (кг, см, % і т. ін.).
Якщо потрібно порівняти ступінь мінливості різнойменних ознак: надій (кг) і вміст жиру в молоці (%) показник сигми переходить у відносну величину. Для цього обраховують коефіцієнт варіації (Cv), який представляє собою відношення величини сигми до середньоквадратичної, виражене у відсотках, –
Cv= (s: Х) х 100 %
Варіансу (s2) отримують підведенням середньоквадратичного відхилення до квадрату. Показник варіанси використовується в генетичному аналізі, коли вимагається розкласти фенотипову мінливість на складові частини: мінливість, обумовлену різноманітністю генотипів особин сукупності, і мінливість, обумовлену впливом різних факторів середовища. Співвідношення варіанси фенотипової (sP2), генотипової (sG2) і паратипової (sE2) можна записати так:
sP2=sG2+sE2
Величини мінливості показника s та s2 мають велике значення в генетичному аналізі популяцій, а також у селекції тварин. Висока мінливість ознаки створює сприятливі умови для селекції, підвищуючи її ефективність.
Для характеристики окремо взятої особини користуються показником нормального відхилення t. Для цього визначають відхилення величини ознаки особин (Х) від середньоарифметичної обстеженої групи (Х). При цьому отримують різницю (Х–Х) і ділять її на величину s, тобто ознаку даної особини виражають у частках сигми:
t = (Х–Х): s
3.6. Показники зв’язку між ознаками
Біометрія дозволяє вивчити зв’язок між варіюючими ознаками, визначити його величину і напрямок. Коефіцієнти, які дозволяють зробити аналіз зв’язків, мають велике практичне значення. Наприклад, важливо встановити, чи великий зв’язок між величиною надою і вмістом жиру у молоці корів, як змінюється рівень жирномолочності при збільшенні надою, який зв’язок між настригом, товщиною і густиною шерсті і т.ін.
Біометричними показниками зв’язку служать коефіцієнти кореляції (r), коефіцієнти регресії (Rxy) і ін. Кореляції потрібно відрізняти від так званих функціональних зв’язків, які характеризують фізичні, хімічні процеси або математичні показники. Функціональні зв’язки характеризуються тим, що при зміні одного показника на певну величину інший показник також змінюється на певну величину. До таких зв’язків відносять, наприклад, зміну довжини кола при зміні величини радіуса і т. ін.
Корелятивні зв’язки відрізняються тим, що при зміні однієї ознаки інша, пов’язана з нею, може мати варіюючі величини у особин даної сукупності. Так, підвищення поживності раціону групи корів на 1 КО буде супроводжуватись у одних особин збільшенням надою на 1,5 кг, у інших – на 0,5 кг, а надої деяких корів групи можуть навіть зменшуватись.
Практичне значення кореляції при вивченні спадковості тварин важливе. Вони дозволяють визначити частку впливу спадковості батька і матері на генотип і фенотип нащадків. Ці коефіцієнти використовують для прогнозування продуктивності даної тварини або всього стада, породи. При виявленні кореляції між ознаками можна проводити опосередковану селекцію. Відбираючи особин за одним яким-небудь бажаним показником, опосередковано здійснюється відбір за іншим показником, корелятивно зв’язаним з основним.
3.7. Статистичні помилки
Вибіркова сукупність є частиною генеральної сукупності, тому одержані при її дослідженні статистичні параметри можуть дещо відрізнятися від величин, характерних для генеральної сукупності. Таке розходження визначають за допомогою статистичних помилок. Для усунення розходження між параметрами генеральної і вибіркової сукупностей вводять поправки на ці параметри у вигляді так званих статистичних помилок (m*): mx, mr, mcv та ін. Знаючи величину статистичної помилки, встановлюють, чи правильно величина параметру вибіркової сукупності відображає величину такого ж параметра генеральної сукупності, тобто визначають статистичну достовірність, або критерії достовірності вибіркових параметрів (t).
Таким чином, обробка масових матеріалів за допомогою біометрії дозволяє правильніше оцінити і охарактеризувати генеральну сукупність тварин за показниками, які вивчаються.
Довгострокові селекційні програми можуть бути здійсненні у тому випадку, якщо розроблення показників стада або породи буде основуватись на використанні генетичних і біометричних параметрів, які характеризують селекційні ознаки.
3.8. Організація масового матеріалу для біометричної обробки
Зоотехнічна і ветеринарна первинна документація дає величезний інформаційний матеріал, який дозволяє всесторонньо охарактеризувати продуктивні і племінні якості стаді, а також економічні показника галузі.
Найпоширенішою формою узагальнення і упорядкування первинних даних є складання варіаційний рядів і корелятивних решіток. Цей спосіб використовується в якості необхідної частини зведених звітів по бонітуванню тварин.
7. Дисперсійний аналіз
Дисперсійний аналіз, розроблений Р. Фішером, дозволяє встановити силу впливу різноманітних факторів на варіювання ознаки, яка вивчається. Зміна ознаки особин даної сукупності виникає під дією багатьох факторів, одні з них можуть знижувати, а інші підвищувати її рівень. При цьому, індивіди, які складають сукупність, неоднаково реагують на весь комплекс зовнішніх умов. Різний ступінь впливу і неоднакова реакція тварин на зовнішні фактори викликає варіювання будь-якої ознаки, що призводить до формування її фенотипової мінливості. Наприклад, фенотипова мінливість надою корів обумовлена спадковістю кожної тварин, рівнем і типом годівлі, елементами технології доїння і утримання віком і т.ін.
Проводячи дисперсійний аналіз, можна встановити, яка частка фенотипової мінливості обумовлена спадковістю, а яка – впливом зовнішніх факторів. Це дуже важливо знати селекціонерам, бо що чим вища генетична частка впливу на фенотип, тим ефективнішою буде селекція.
Лекція 9. МУТАЦІІ ТА МУТАГЕНЕЗ
План:
1. Мутації та їх класифікація
2. Поняття про мутагенез
3. Мутагенні фактори та їх класифікація
4. Штучний мутагенез
1. Мутації та їх класифікація
Поняття “мутація” в генетику ввів Г. до Фріз у роботах 1901-1903 років.
Мутація – разова стрибкоподібна зміна спадкової ознаки у результаті зміни спадкового фактора, який її визначає.
До робіт Г. де Фріза вже були відомі випадки зміни спадковості у рослин і тварин. Це відмічав ще Ч. Дарвін. Проте саме Г. де Фріз сформулював теорію мутацій, яка до теперішнього часу зазнала змін, але у головних питаннях залишалась незмінною і відіграла позитивну роль у розвитку генетичної теорії.
Класифікація мутацій (різні принципи класифікацій мутацій, в основу яких покладені різні принципи).
1. Класифікація мутацій за локалізацією у геномі
1.1. Генні мутації − спонтанні (довільні) або індуковані (штучні) спадкові зміни молекулярної структури гена з утворенням нових алелей.
Різновиди генних мутацій:
Ø транзиція – заміщення одного пурину або одного піримідину іншим (А«Г, Т«Ц);
Ø трансверсія – заміщення пурину піримідином і, навпаки (А«Т, Ц«Г);
Ø делеція – втрата одного нуклеотиду (нуклеотидної пари);
Ø інсерція – вставка одного нуклеотиду (нуклеотидної пари).
Механізм виникнення:
Ген у структурному відношенні – це ділянка молекули ДНК, яка складається із нуклеотидів. Генетична інформація у ньому записана триплетами: кожні три нуклеотиди, що розміщені підряд, кодують певну амінокислоту. Тому від послідовностей розміщення нуклеотидів у кожному триплеті і розміщення самих триплетів залежить послідовність амінокислот у синтезованій молекулі білка, тобто тип білка і його функція. Якщо в одному триплеті (кількість триплетів у середньому гені сягає 500) випаде хоч один нуклеотид, то відбудеться зміщення рамки зчитування інформації. Отже, послідовність амінокислот у майбутній молекулі білка буде іншою, тобто утвориться інший білок. Функція білка також зміниться повністю або частково.
Наслідки генних мутацій:
Транзиції і трансверсії викликають, на перший погляд, незначні зміни генів, проте наслідки їх можуть бути тяжкими. Прикладом трансверсії є утворення мутантного гемоглобіну S, замість нормального, А, у людини. Мутація призвела до відмінності між двома типами гемоглобіну лише за однією амінокислотою:
| Гемоглобін А – пролін-глютамін-глютамін-лізин Гемоглобін S – пролін-валін-глютамін-лізин |
Але, не дивлячись на таку незначну різницю, гемоглобін S набув інших властивостей, які відбиваються на життєдіяльності організму. Гомозиготні особини за геном гемоглобіну S мають еритроцити серпоподібної форми (форми півмісяця) і гинуть внаслідок особливого типу анемії (серпоподібноклітинної).
Делеція та інсерція призводить до менших або більших наслідків залежно від місця їх виникнення. Якщо це відбувається, наприклад, в останньому триплеті гена, то він незначно змінить свою структуру, а якщо у першому, то структура його зміниться повністю внаслідок зміщення рамки зчитування. Безумовно, що відповідно зміниться структура і функція тих білків, які кодуються мутантними генами.
Поширеність генних мутацій у природі:
Порівняно з іншими мутаціями дуже поширені.
Значення генних мутацій:
Можуть бути корисними, байдужими і шкідливими для організму.
1.2. Хромосомні мутації (аберації) − зміна не одного гена, а двох або декількох, що призводить до зміни форми хромосом.
Різновиди аберацій:
Ø нестача – втрата кінцевого фрагмента;
Ø делеція – втрата середнього фрагмента;
Ø дуплікація – подвоєння будь-якого фрагмента хромосоми;
Ø інверсія – поворот фрагмента хромосоми на 180°;
Ø інсерція (транслокація інтрахромосомна) – переміщення фрагменту тієї самої хромосоми в інше положення;
Ø транслокація інтерхромосомна – вклинення фрагментів однієї хромосоми в іншу.
Поширеність аберацій у природі:
На теперішній час вивчені деякі хромосомні мутації у людини та різних видів сільськогосподарських тварин. Наприклад, у великої рогатої худоби відома інтерхромосомна транслокація 1/29, у свиней 4/14 та ін.
Значення аберацій:
За генетичними наслідками аберації неоднозначні. Так, виникнення нестач, делецій і дуплікацій пов'язане з порушенням генного балансу, що може призвести до різних змін за фенотипом і навіть до летальних наслідків. Водночас інверсії й інтрахромосомні транслокації на сильні фенотипові зміни майже не впливають, але, як правило, зміна положення гена не проходить безслідно для організму. Наприклад, дія структурних генів, які розміщені поряд і беруть участь в одній і тій самій ланці синтезу, при переміщенні порушується. Оскільки високочутливих біохімічних методів вияву порушень синтезу окремих речовин немає, то можна тільки здогадуватись про наявність ще не відомих або відомих, але не пізнаних (у причинному відношенні) молекулярних хвороб.
1.3. Геномні (плоїдні) мутації − збільшення або зменшенням кількості хромосом у каріотипі організму. Бувають:
1.3.1. Диплоїдні − збільшення кількості хромосом з одинарного до подвійного (наприклад, збільшення кількості хромосом з одинарного (1n) до подвійного (2n) у природних гаплоїдів (гаметофіт вищих рослин, трутні медоносної бджоли);
1.3.2. Еуплоїдні (гаплоїдні) – зменшення хромосомного набору з подвійного до одинарного (гаплоїдні спорофіти рослин виживають, але мають менші розміри та знижену життєздатність; гаплоїдних мутацій тварин у тварин не зафіксовано, тому що їх розвиток дуже швидко припиняється на перших стадіях дробіння з утворення виродків);
1.3.3. Поліплоїдні (триплоїдні, тетраплоїдні, пентаплоїдні, гексаплоїдні і т. д.) – кратне збільшення кількості хромосом на гаплоїдниї хромосомний набір.
Причини поліплоїдії:
Ø порушення мейотичного поділу з утворенням диплоїдних гамет;
Ø поліандрія (поліспермія) – запліднення яйцеклітини двома і більше сперматозоїдами;
Ø полігінія – злиття при заплідненні кількох пронуклеусів (ядер яйцеклітин) і сперматозоїда;
Ø анеугамія − гібридизація (схрещування) особин з різною плоїдністю;
Ø затримка першого поділу зиготи – дробіння, за якого відбувається подвоєння хромосом, а поділу ядра і цитоплазми не відбувається.
Різновиди поліплоїдії:
Ø парна – ортоплоїдія (4n);
Ø непарна – анортоплоїдія (3n).
Поширеність поліплоїдії у природі:
На Крайній Півночі і високо в горах поліплоїдні форми рослин становлять 80 %.
Дуже часто поліплоїдія зустрічається у безхребетних тварин (мокриці, черви та інші гермафродитні й партеногенетичні види).
Поліплоїдні форми вищих тварин у більшості випадків нежиттєздатні (їх виявлено лише серед абортованих плодів домашніх тварин і людини; О. О. Іванова (1974) відмічає випадок народження триплоїдного хлопчика, який, мав значні аномалії і прожив короткий період.
Значення поліплоїдії у природі:
Поліплоїдія зумовлює стійкість рослин до несприятливих умов середовища.
Значення поліплоїдії для людини:
Дуже широко використовують у селекції рослин, для створення форм, які характеризуються дуже високою продуктивність, порівняно із звичайними. Тому переважна більшість сільськогосподарських рослин – поліплоїдні (картопля, цукрові буряки, суниця та ін.). Розрізняють автополіплоїдію – поліплоїдію у межах одного виду рослин та аллополіплоїдію (амфіполіплоїдію) – міжвидові поліпоїди (гібрид капусти і редьки (рафанобрасіка), пирію і жита (тритикале).
1.3.4. Анеуплоїдні (гетероплоїдні ) – збільшення або зменшення кількості хромосом на одну або дві хромосоми у кожній парі хромосом.
Причина анеуплодії:
Ø порушення мейозу, при якому гомологічні хромосоми у профазі І редукційного поділу або зовсім не кон’югують, або кон’югують частково, що призводить надалі до нерозходження хромосом, тобто вони розподіляються між полюсами випадково. Так, обидві хромосоми можуть відійти до одного полюса, тоді одна з клітин матиме одну зайву хромосому, а в іншій однієї хромосоми не вистачатиме.
Різновиди анеуплодії:
1) Залежно від кількості хромосом у нормі, зайвих або відсутніх у соматичних клітинах:
| Ø диплоїдія (дисомія) | 2n (норма) |
| Ø моносомія | 2n – 1 |
| Ø нулісомія | 2n – 2 |
| Ø подвійна моносомія | 2n – 1 – 1 |
| Ø трисомія | 2n + 1 |
| Ø тетрасомія | 2n + 2 |
| Ø подвійна трисомія | 2n + 2 + 1 |
| Ø подвійна тетрасомія | 2n + 2 + 2 |
2) Залежно від типу хромосом:
Ø аутосомну – порушення нормального співвідношення аутосом;
Ø гоносомна − порушення нормального співвідношення статевих хромосом.
Поширеність анеуплодії у природі:
У людини відомі випадки аутосомних анеуплодій, які обумовлюють синдром Дауна (трисомія-21), Патау (трисомія-13 (14 чи 15), Едвардса (трисомія-16 (17 чи 18). Інші анеуплоїдні форми аутосомного походження у людей не зустрічаються, тому що гинуть в період ембріонального розвитку внаслідок сильного зниження життєздатності. Доказом цього є виявлені анеуплоїди у довільно абортованих плодах. Відомостей про аутосомних анеуплоїдів у тварин зовсім мало. Мабуть це теж пояснюється довільно абортованими нежиттєздатними плодами.
У людини відомі випадки гоносомних анеуплодій, які обумовлюють синдром Клайнфельтера (ХХУ), Шерешевського-Тернера (Х0), трисомії за Х-хромосомою (XXX), полісомії за У-хромосомою (ХУУ).
У великої рогатої худоби, свиней, собак, котів відомий синдром Клайнфельтера (ХХУ).
Значення анеуплодій:
Анеуплоїдні організми мають знижену життєздатність, оскільки кожна хромосома має свої конкретні гени, то їх втрата або надлишок призводить до різних аномалій чи захворювання.
Інші принципи класифікації мутацій
| Назва | Визначення | Приклад | Значення | ||
| Класифікація мутацій в еволюційному аспекті | |||||
| ретрогресивні | Призводять до втрати ознаки. | паразитичні черви, у яких втрачені або недорозвинені деякі органи; сліпаки, у яких відсутній зір; коні, у яких однопала кінцівка. | Ці мутації не можна розглядати, як процес виродження. Вони можуть відігравати істотну пристосувальну роль. Не дивлячись на втрату деяких органів види успішно процвітають. | ||
| дегресивні | Перехід гена у процесі послідовних актів мутування з одного стану в інший. | альбінізм і забарвлене волосся у тварин; рогатість і комолість. | Відіграють пристосувальну роль. | ||
| прогресивні | Призводять до утворення нової ознаки. | набуття хлорофілу рослинами, хребта хордовими | Можуть дати початок новим видам. | ||
| Класифікація мутацій за характером дії гена (функціональною дією гена) була запропонована Г. Менделем у 1927 році. | |||||
| аморфні (неактивні або нейтральні) | Призводять до утворення функціонально неактивного генного продукту, у результаті чого ознака, яка ним контролюється, не розвивається. | альбінізм, беззубість, відсутність кінцівок, безхвостість | Часто така мутація летальна у гомозиготному стані гена, з чим пов’язана ембріональна смертність тварин. Однак іноді такі мутації є корисними, як, наприклад, біле хутро у полярного ведмедя. Втрата інстинкту насиджування у домашніх курей є корисною для людини. | ||
| гіпоморфні | Послаблюють дію нормального гена і призводять до послаблення вираження ознаки у порівнянні з вихідним типом. | різні прояви карликовості, недорозвинення органів, послаблення забарвлення волосся тощо | Частота таких мутацій найбільша, порівняно з іншими, тому що будь-яка мутація у більшості випадків здатна погіршити ознаку, ніж змінити її у кращу сторону.
У гомозиготному стані такі мутації бувають летальні.
Часто такі зумовлюють різноманітність ознаки у популяції. Цей тип мутацій має важливе значення у селекції хутрових звірів для ство Date: 2015-09-02; view: 2509; Нарушение авторских прав |