Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Цитогенетика як наука
|
|
До початку XX століття завдяки спостереженням, проведеним на різноманітних рослинних і тваринних клітинах, були встановлені багато деталей їхньої будови, відкритий мітоз, вивчений хід оогенезу, сперматогенезу і запліднення. Зіставлення цих великих цитологічних відкриттів з результатами гібридологічних дослідів привело до нового більш глибокого розуміння природи спадковості і послужило могутнім поштовхом до швидкого подальшого прогресу генетики.
Весь період розвитку генетики від часу відкриття законів Г.Менделем до 50-х років ХХ сторіччя (до вивчення генетичних явищ на молекулярному рівні) був ознаменований синтезом гібридологічних і цитологічних даних, що виразилися у створенні цитогенетики – науки про цитологічні основи спадковості.
Плодом тісного зближення генетики і цитології стала хромосомна теорія спадковості – найважливіше узагальнення, яке багато в чому визначило подальший розвиток генетики, і справило великий вплив на багато інших галузей біології.
Цитологічні основи спадковості були з'ясовані дослідженнями, проведеними головним чином на багатоклітинних організмах, які усі відносяться до еукаріот, тобто мають морфологічно відособлене від цитоплазми клітинне ядро. Ці дослідження пролили світло на внутрішньоклітинні механізми генетичних процесів, а висновки з них надалі послужили передумовою для вивчення генетичних явищ на молекулярному рівні, що було у великій мірі зв'язано з розгортанням робіт з генетики прокаріотів, тобто одноклітинних організмів, що не мають відособленого ядра, головним чином бактерій, а також по генетиці вірусів, що представляють неклітинну форму життя.
Цитогенетика використовує однойменний метод – цитогенетичний – який дозволяє вивчити особливості передачі генетичної інформації на клітинному рівні. У його завдання входить вивчення будови і функції клітинних структур, які приймають участь у передачі спадковості. Частиною цього методу є каріологічний аналіз – дослідження кількості, розмірів, форми і фізико-хімічних властивостей хромосом, як основних носіїв спадковості.
2. Клітина як матеріальна основа спадковості
Після відкриття мітозу, оогенезу, сперматогенезу і запліднення німецькі учені А. Вейсман, О. Гертвіг і Е.Страсбургер висунули гіпотезу, що спадкові ознаки передаються ядром. Цю думку було підкріплено численними експериментально-цитологічними й експериментально-ембріологічними роботами.
Одна з перших робіт такого роду була проведена німецьким ембріологом К. Гербстом на морських їжаках: у своїх досвідах він використовував морських їжаків двох різних родів, личинки яких дуже відрізняються будовою кістяка і деяких інших ознак. При цьому яйця він брав від одного роду, а спермії – іншого. У личинок, які виходили, та половина тіла, клітини якої мали ядра, що походять тільки від ядра яйця, мала винятково материнські ознаки, інша ж половина, клітини якої містили гібридні ядра, характеризувалася явно проміжною будовою, що суміщала ознаки обох родин морських їжаків.
Дуже переконливі дані про відносну роль ядра і цитоплазми у спадковості були отримані в СРСР Б. Л. Астауровим у досвідах із шовкопрядами:
| Б.Л. Астауров у незапліднених яйцях шовковичного шовкопряда тепловим шоком і опроміненнями рентгенівськими променями руйнував ядра. Потім такі яйця осіменяв. При цьому ядра двох сперміїв, які проникли в яйце, зливалися і утворювали ядро зиготи. Розвинені з таких андрогенних зигот особини шовкопряда завжди були самцями і точно повторюють фенотип своїх батьків. Це було особливо помітно, якщо яйця належали самкам однієї породи, а спермії – самцям іншої. Подібні досвіди Астаурову вдалося провести, руйнуючи зазначеним способом ядра в яйцях іншого виду шовкопрядів – мандаринового – і осіменяючи їх сперміями шовковичного шовкопряда. | 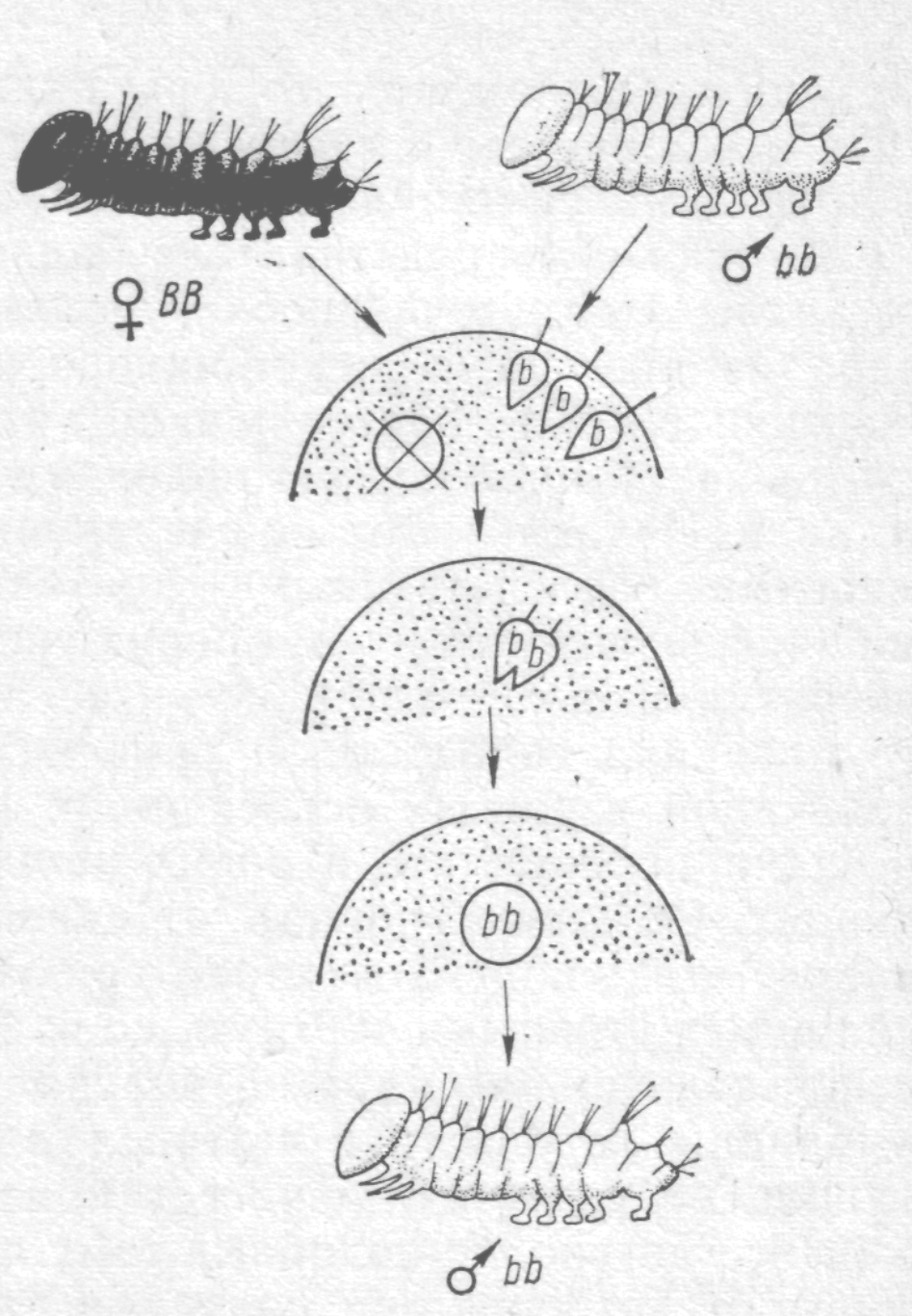 Андрогенез у шовковичного шовкопряда (за Астауровим)
Андрогенез у шовковичного шовкопряда (за Астауровим)
|
Зручним об'єктом для дослідження ролі ядра служать великі прісноводні амеби:
| Більшість дослідників прийшли до висновку, що за всіма ознаками амеби з пересадженими ядрами ставали цілком схожими на амеб того штаму, від якого вони одержали ядро, і що вони передавали ці ознаки без зміни наступним поколінням (в одному випадку це було простежено більш ніж у 300 поколіннях амеби з пересадженим ядром). | 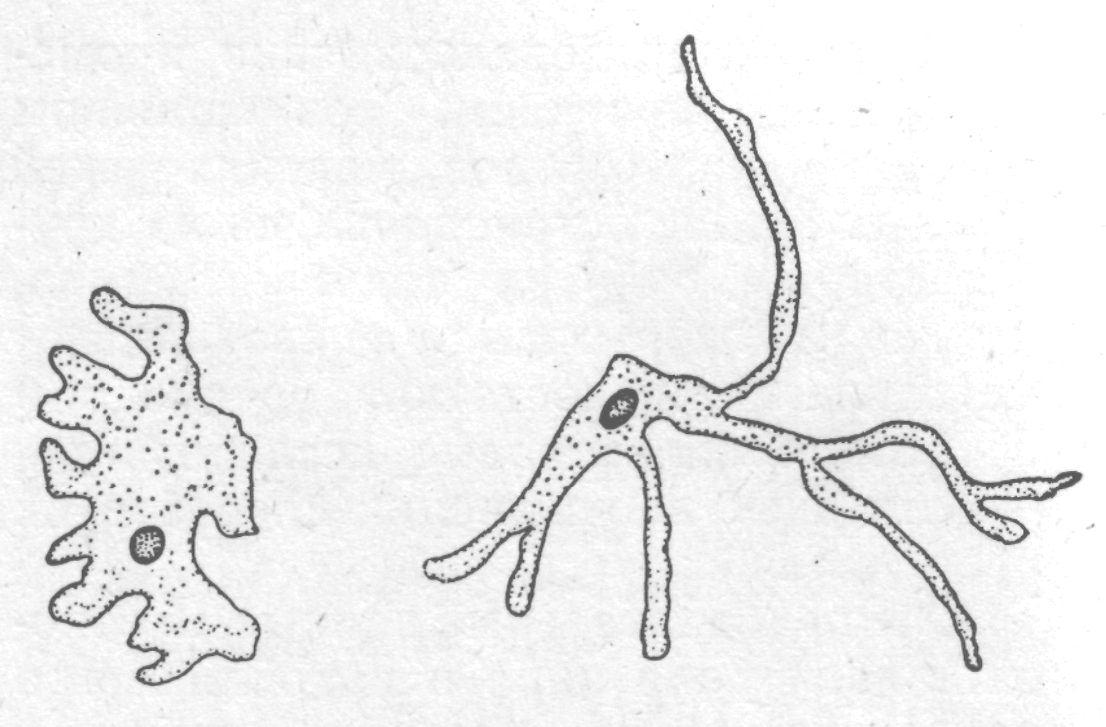 Спадкова різниця у формі тіла амеби прісноводної
Спадкова різниця у формі тіла амеби прісноводної
|
Про провідну роль ядра у спадковості у рослин служить дослід на морській водорості ацетабулярії:
| Форма шапочки дуже різна в різних видів ацетабулярій. Якщо зрощувати відрізане стебельце одного виду з ризоїдом іншого, то шапочка, яка регенерує на стебельці має форму, властиву виду, якому належить ядро. | 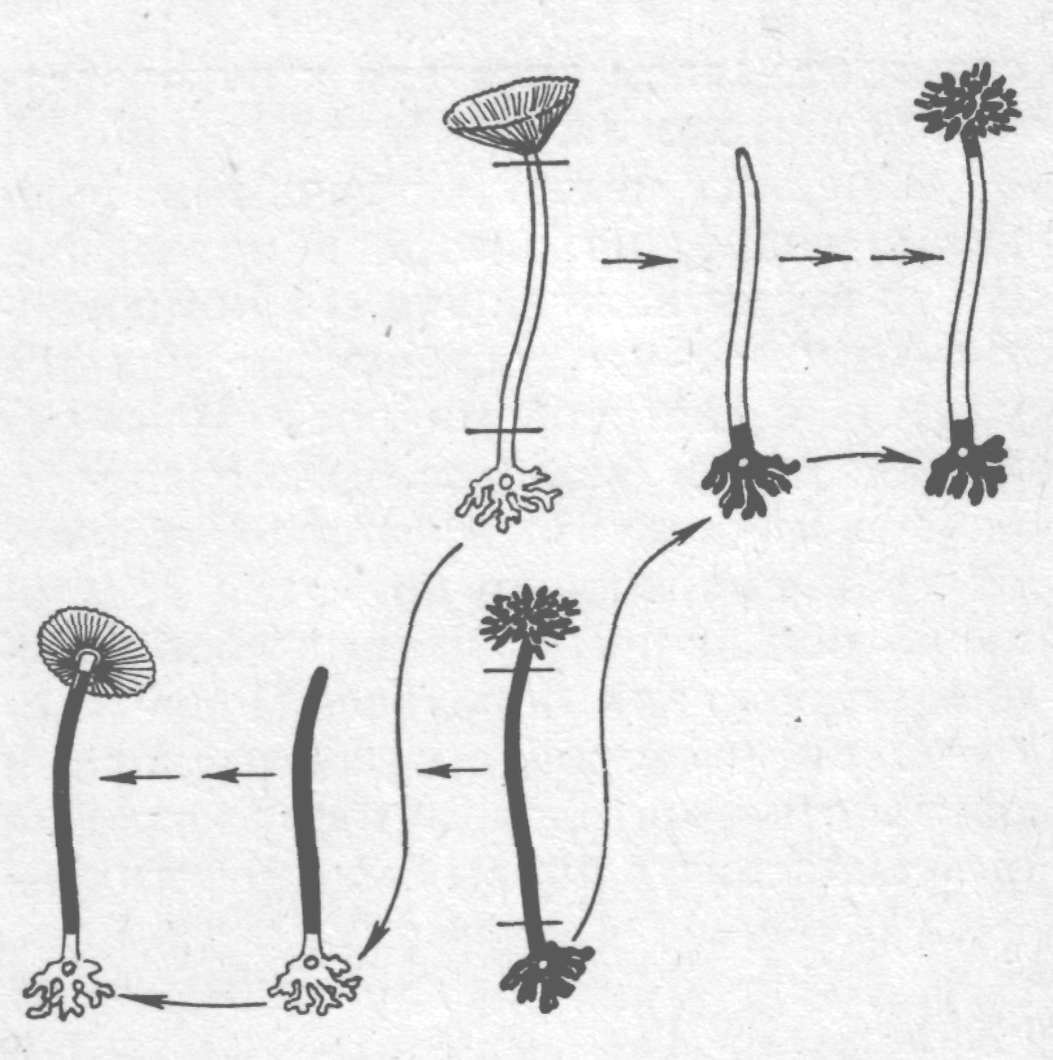 Регенерація при перехресному зрощенні в ацетабулярії
Регенерація при перехресному зрощенні в ацетабулярії
|
Найбільш прямі свідчення про відносне значення ядра і цитоплазми в спадковості вищих тварин дали досліди по пересадженню ядер різних земноводним (тритонами, аксолотлями, ропухами, жабами): за допомогою мікроманіпулятора з яйця, яке почало розвиватися, видаляли ядро і замість нього пересаджували в це яйце ядро, взяте з клітки зародка якої-небудь іншої раси того ж чи близького виду. З яєць з пересадженими ядрами виходили ембріони, пуголовки і дорослі жаби, за всіма ознаках завжди цілком подібні до того підвиду, від якого вони одержали ядро, і не виявляли жодної ознаки іншого підвиду, від якого яйця одержали цитоплазму.
У той же час потрібно відзначити, що відомо і деякі достовірні випадки успадкування ознак через цитоплазму, а точніше її органоїди. Доказами цього є, наприклад, пластидна спадковість.
Досвіди по пересадженню ядер свідчать про те, що у різних за своїм систематичним положенням організмів найрізноманітніші спадкові ознаки визначаються клітинним ядром.
Отже, клітина є матеріальним носієм спадковості. Провідну роль у передачі спадкової інформації від клітин до клітини відіграє ядро, яке тісно пов’язана з цитоплазмою. Воно перебуває у системі зворотного зв’язку з нею. Цитоплазма змінюється під впливом генів, які зосереджені у ядрі, але у результаті цього змінюється і цитоплазма, тобто середовище у якому діють гени. Тому, єдність ядра і цитоплазми нерозривна і постійна.
3. Хімічний склад і морфологічна будова хромосом
3.1. Докази ролі хромосом у спадковості
Хромосоми є безпосередніми носіями спадкової інформації у ядрі. Багато зусиль вчених пішло на підтвердження провідної ролі хромосом у спадковості.
Є прямі і опосередковані докази ролі хромосом у спадковості.
Опосередковані докази ролі хромосом у спадковості:
ü хромосоми є постійними органоїдами ядра;
ü на відміну від інших клітинних органоїдів, відрізняються постійністю числа і форми у певних видів;
ü клітинне ділення і весь процес статевого розмноження сприяють збереженню постійності хромосом у дочірніх клітинах і у наступних поколіннях.
Прямі докази ролі хромосом у спадковості:
ü дослід Блекслі:
| вивчаючи рослину дурману, яка має 12 пар хромосом, встановив, що при появі у каріотипі зайвої хромосоми змінювався ряд ознак даної рослини. За зовнішнім виглядом навіть можна було встановити, яких саме хромосом стало 3. На малюнку показано зміну коробочок дурману залежно від того, у якій парі хромосома зайва. |  Коробочки дурману:
1 – нормальної диплоїдної рослини з 24 (2n) хромосомами; 2-13 – рослини, які мають 24+1 хромосоми (трисоміки)
Коробочки дурману:
1 – нормальної диплоїдної рослини з 24 (2n) хромосомами; 2-13 – рослини, які мають 24+1 хромосоми (трисоміки)
|
На теперішній час хімічний склад та морфологічна будова хромосом добре вивчена.
3.2. Хімічний склад і морфологічна будова хромосом
Речовину, яка утворює хромосоми, називають хроматином, у склад якого входить одна молекула ДНК та основні білки-гістони.
Спостерігати і вивчати морфологічну будову хромосоми можна лише у стані їх спіралізації, яка відбувається під час поділу клітини (у метафазі мітозу).
Хромосоми існують у ядрі протягом всього життя клітини, але залежно від стадії мають різну форму і величину.
В період інтерфази (відносного періоду спокою клітини) – це деспіралізовані нитки ДНК, які досягають декількох сантиметрів у довжину і мають товщину 20-30 нм. У такому стані їх не можна побачити у мікроскоп.
У період поділу клітини хромосоми спіралізуються і стають помітні у мікроскоп.
У період мітозу хромосоми – це компактні тільця характерної форми, довжиною 1,5-10 мкм і товщиною 0,2-2 мкм. Для того, щоб хромосома могла прийняти такі розміри, вона повинна укоротитись в 10 тисяч разів, тобто сконденсуватися.
У цей час величезна молекула вигинається, утворюючи дрібні, щільно складені петлі, скріплені негістоновими білками, які називаються хромомерами. Вони розміщені на хромосомі через відповідні проміжки. У склад одного хромомера входить 50-70 тисяч пар нуклеотидів, що відповідає розміру одного гена.
Подальша конденсація призводить до утворення хромонеми – нитки, яка утворюється за рахунок зближення хромомерів. Хромонеми вже помітні у світловому мікроскопі.
Хромонема конденсується і утворює хроматиду. Хромосома має одну чи дві хроматиди залежно від фази ділення.
Мітотична хромосома оточена рибонуклеопротеїдами, білками і іншими речовинами. Це матрикс хромосоми. З матриксом хромосоми передають дочірніми клітинам матеріал, необхідний для формування ядерця і початкового етапу білкового синтезу в молодих клітинах.
Повністю конденсована мітотична хромосома має два плеча, які називають теломерами, між якими є первинна перетяжка, або центромера (кінетохор) – ділянка, до якої приєднуються нитки веретена під час поділу клітини. Деякі хромосоми мають вторинну перетяжку. Деякі вторинні перетяжки пов’язані з формуванням ядерця, тому їх називається ядерцевим організатором. Ця ділянка хромосоми має складну структуру і відповідає за синтез рибосомної РНК. Іноді вторинна перетяжка може бути дуже довга, а іноді вона відділяє від основного тіла хромосоми невелику ділянку, яка називається супутником.
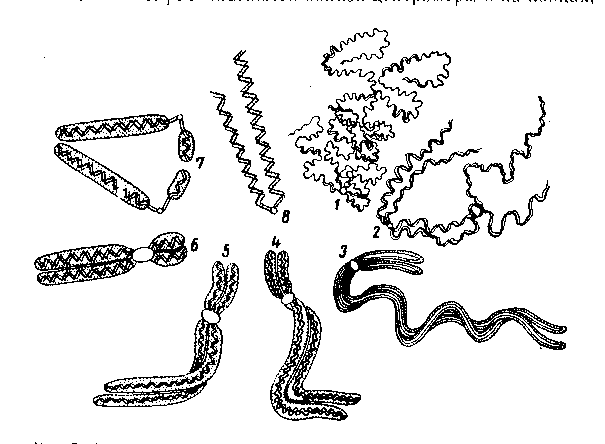 Схема спіралізації хромонем в мітотичному циклі:
1 – інтерфаза, хромономи слабоспіралізонані; 2, 3, 4 – профаза, посилення спіралізації хромонем, утворення двох хроматид; 5 – прометафаза, поява чотирьох напів хроматид; 6 – метафаза, максимальна спіралізація, помітні велика і мала спіраль; 7 – анафаза; 8 – телофаза (одна з дочірніх хромосом), де спіралізація хромонем
Схема спіралізації хромонем в мітотичному циклі:
1 – інтерфаза, хромономи слабоспіралізонані; 2, 3, 4 – профаза, посилення спіралізації хромонем, утворення двох хроматид; 5 – прометафаза, поява чотирьох напів хроматид; 6 – метафаза, максимальна спіралізація, помітні велика і мала спіраль; 7 – анафаза; 8 – телофаза (одна з дочірніх хромосом), де спіралізація хромонем
| 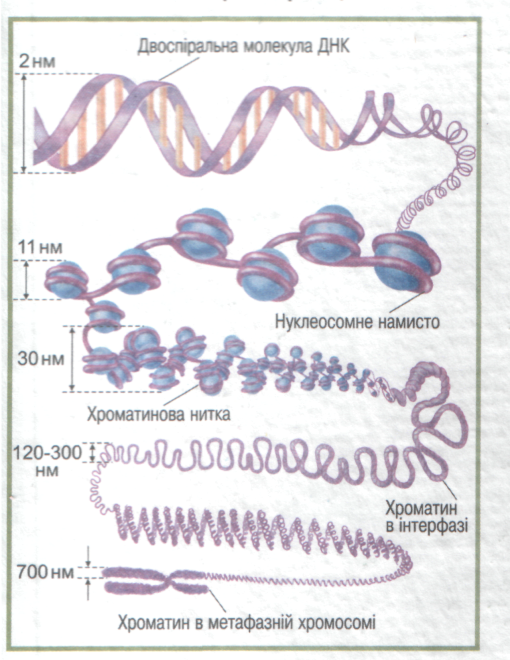 Схема компактизації молекули ДНК
з утворенням метафазної хромосоми
Схема компактизації молекули ДНК
з утворенням метафазної хромосоми
|
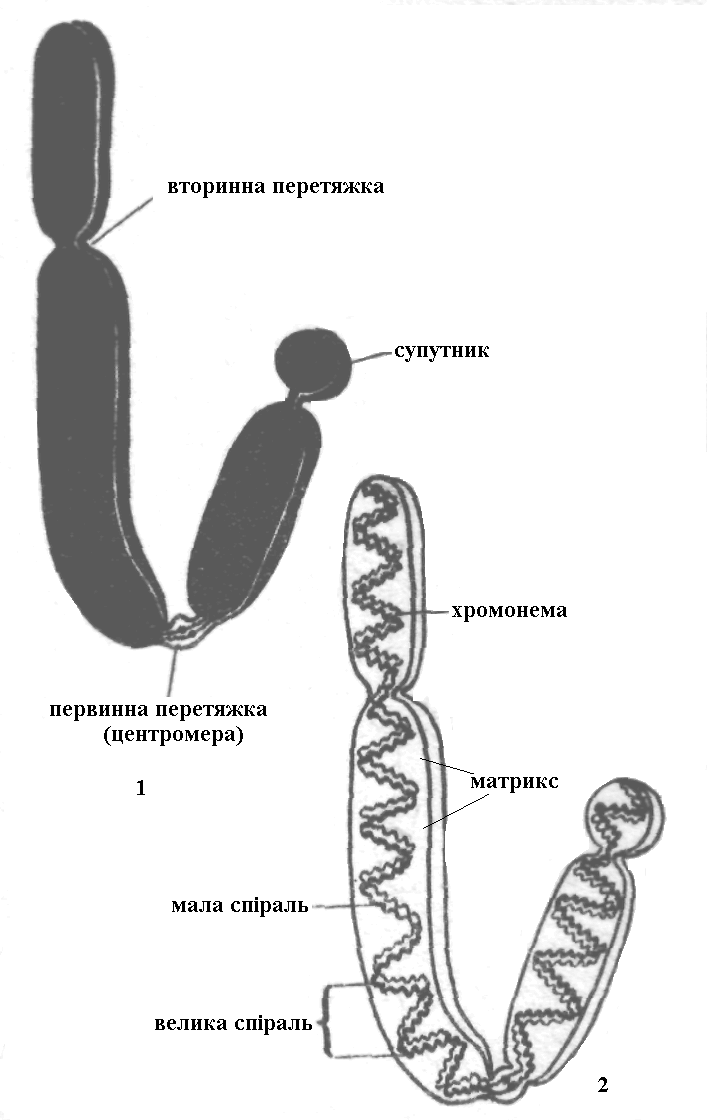 Схема будови метафазної хромосоми:
1 – морфологія; 2 – внутрішня структура хроматиди
Схема будови метафазної хромосоми:
1 – морфологія; 2 – внутрішня структура хроматиди
| 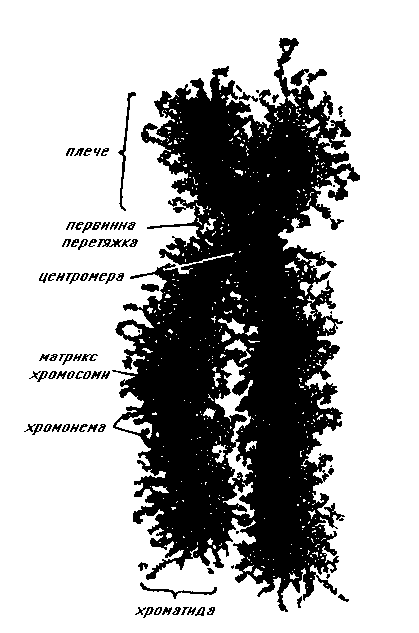 Схема будови метафазної хромосоми
Схема будови метафазної хромосоми
|
Кожна хромосома має характерну постійну форму і розміри. Форма хромосом залежить від місця розташування центром ери:
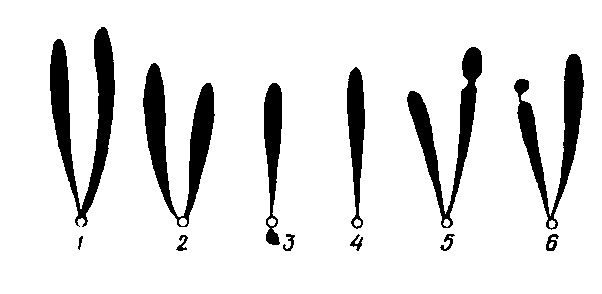
Типи метафазних хромосом:
1 – метацентрична (рівноплеча); 2 – субметацентрична (слабонерівноплеча); 3 – акроцентрична (різконерівноплеча); 4 – телоцентрична (одноплеча); 5 – центрична із вторинною перетяжкою; 6 – супутнична
Для вивчення хромосом використовують спеціальне фарбування, при якому різні ділянки хромосом забарвлюються з різною інтенсивністю, утворюючи ділянки:
ü гетерохроматинові – інтенсивно забарвлені;
ü еухроматинові – слабо забарвлені.
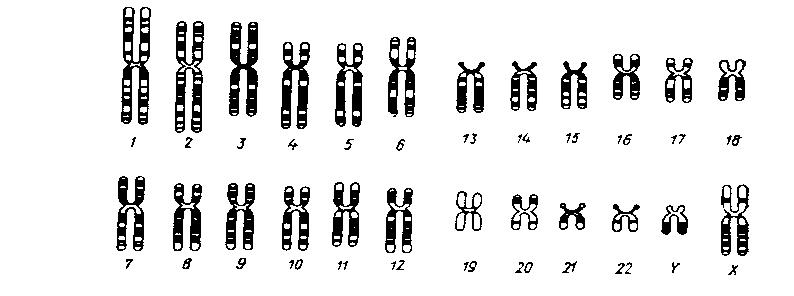
Диференційоване фарбування хромосом людини (схематично)
Завдяки даному методу вдається ідентифікувати гомологічні хромосоми.
Каріотип – сукупність хромосом певного розміру і форми у соматичних клітинах певного виду організмів. Кількість хромосом у каріотипі певного виду не залежить від рівня його організації і систематичного положення. Найчастіше зустрічаються каріотипи, які нараховують від 10 до 60 хромосом, але відомі і з двома, а також десятками і навіть сотнями хромосом.
У всіх статевих клітинах організмів одинарний, або гаплоїдний набір хромосом (1n). У результаті злиття двох гаплоїдних статевих клітин утворюються соматичні клітини з подвійним, або диплоїдним набором хромосом (2n).
Диплоїдний набір деяких тварин
| Організм | 2n | Організм | 2n |
| Людина – Homo sapiens | Ропуха – Bufo bufo | ||
| Шимпанзе – Pan troglodytes | Квакша – Hyla arborea | ||
| Макака резус – Macacus rhesus | Тритон – Triturus vulgaris | ||
| Кінь – Equus caballus | Окунь – Perca fluviatilis | ||
| Кінь Пржевальського – Equus przewalskii | Сазан – Cyprinus carpio | ||
| Осел – Equus asinus | Карась – Carassius arrutus | 94-100 | |
| Свиня домашня – Sus scrofa | Муха хатня – Musca domestica | ||
| Кабан дикий – Sus scrofa | Мушка плодова – Drosophila melanogaster | ||
| Вівця домашня – Ovis anies | Комар-пискун – Culex pipiens | ||
| Корова – Bos taurus | Бджола медоносна – Apis mellifera самець самка | ||
| Коза домашня – Capra hircus | Шовкопряд шовковичний – Bombyx morі | ||
| Кішка – Felis catus | Попелиця оранжерейна (персикова) – Muzus persicae | ||
| Лисиця – Vulpes vulpes | Воша головна – Pediculus capitis | ||
| Собака домашній – Canis familiaris | Тарган рудий – Blattella germanica самець самка | ||
| Миша хатня – Mus musculus | Сарана азіатська – Locusta migratoria | ||
| Пацюк сірий – Rattus norvegicus | Кліщ собачий – Ixodes ricinus | ||
| Кріль – Oryctolagus cuniculus | Рак річковий – Astacus fluviatilis | приблизно 116 | |
| Норка – Mustella visen | Краб – Eupagurus ochotensis | приблизно 254 | |
| Курка домашня – Callus domesticus | Аскарида кінська – scaris megalocephata | 2, 4 | |
| Індик – Melleagris gallopavo | Виноградний слимак – Helix pomatia | 24, 48 | |
| Голуб – Columba livia | Черв’як дощовий – Lumbricus terrestris | ||
| Качка кряква – Anus platyrhincha | Планарія біла – Dugesia gonocephala | 16-32 | |
| Ящірка прудка – Lacerta agilis | Гідра прісноводна – Hydra oligactis | ||
| Жаба озерна – Rana ridibunda | Малярійний плазмодій – Plasmodium malariae |

Каріотипи соматичних клітин деяких тварин і рослин в метафазі:
1 – лісової жаби (26 хромосом); 2 – вівці (54 хромосомы); 3 – людини (46 хромосом); 4 – котячої аскариди (8 хромосом); 5 – комара Anophelus (6 хромосом); 6 – дрозофіли самця и самки (8 хромосом); 7 – жита (14 хромосом); 8 – твердої пшениці (42 хромосомы); 9 – люпину жовтого (48 хромосом); 10 – проса (36 хромосом); 11 – курки (78 хромосом); 12 – великої рогатої худоби (60 хромосом); 13 – коня (66 хромосом)
У диплоїдному наборі розрізняють гомологічні і статеві хромосоми.
Гомологічні хромосоми, або аутосоми – парні хромосоми диплоїдного набору, які мають однакову форму і розмір та розміщенням гетеро- і еухроматинових ділянок. У цій парі одна хромосома – материнська, друга – батьківська.
Статеві хромосоми – це пара негомологічних хромосом диплоїдного набору, за якими розрізняються самці і самки.
4. Цитологічні основи нестатевого розмноження
Нестатево клітини та організми розмножуються шляхом мітозу, амітозу та ендомітозу.
Клітини та організми, які розмножуються мітозом протягом свого існування проходять певний життєвий цикл, який включає інтерфазу та мітоз.
Мітоз – непрямий поділ еукаріотичних клітин, що складається з ділення ядра (каріокінезу) і ділення цитоплазми (цитокінезу). Він супроводжується морфологічною перебудовою ядер і цитоплазми клітини. Це безперервний процес, у якому кожна стадія непомітно переходить в іншу, але для зручності вивчення виділяють чотири фази: профазу, метафазу, анафазу і телофазу.
Амітоз – це прямий поділ клітин без морфологічної перебудови їх ядер і цитоплазми. Він не обов’язково закінчуються поділом цитоплазми, обмежуючись лише поділом ядра. У результаті утворюються багатоядерні клітини. Поділ відбувається шляхом перешнуровування ядра на дві частини: іноді з одного ядра утвориться відразу кілька ядер (фрагментація). Клітина, в якій відбувся амітоз, надалі в нормальний мітотичний цикл не вступає, тому що хромосоми розділяється між дочірніми клітинами нерівномірно.
Амітоз зустрічається майже завжди у клітинах тканин, які віджили себе і приречені до загибелі або у клітинах тканин тимчасово існуючих утворів (клітини плідних оболонок, фолікулярні клітини яєчника). Амітозом діляться клітини і при патологічних процесах – запаленнях, регенерації, злоякісному рості. Тобто амітоз зустрічається у тих випадках, коли необхідне швидке накопичення маси клітин. У цьому полягає його біологічне значення, якщо не брати до уваги злоякісний ріст.
Ендомітоз. Ендомітозом називають такий процес, при якому репродукція хромосом у клітині не супроводжується поділом ядра. Внаслідок цього у клітині відбувається множення числа хромосом, іноді в десятки разів у порівнянні з вихідним. Ендомітоз зустрічається в інтенсивно функціонуючих клітинах різних тканин як рослин, так і тварин. Іноді відтворення хромосом відбувається без збільшення їхнього числа в клітині. Кожна хромосома багаторазово подвоюється, але дочірні хромосоми залишаються зв'язаними між собою. Це явище називається політенією. Воно являє собою окремий випадок ендомітозу. Число ниток у політенній хромосомі може досягати 1000-2000. У цьому випадку утворяться так звані гігантські хромосоми. Явище політенії спостерігається в клітинах ряду диференційованих тканин і зв'язано з особливими функціями ядра клітини.
Життєвий цикл клітини
| Характеристика | Тривалість | |
| Клітинний цикл | Період існування клітини від одного поділу до іншого, який включає інтерфазу і мітоз.
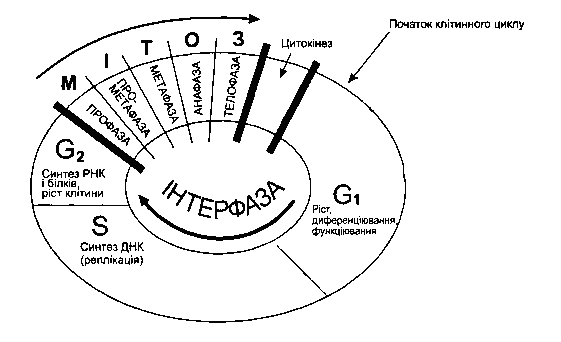 Схема життєвого циклу клітини
Схема життєвого циклу клітини
| Залежить від: · типу тканини (епітеліальні клітини кишечнику – 19 годин, епітелію шкіри – майже 586 годин); · виду організму (у прокаріот – 20-30 хв, в еукаріот – 10-80 год). |
| Інтерфаза (від лат. інтер – між і грец. фазіс – поява) періоди: | Період росту і активного функціонування клітини, який включає постмітотичний, синтетичний та постсинаптичний періоди. | Зазвичай до 90% часу усього клітинного циклу. Співвідношення тривалості фаз залежить · типу тканини (у клітинах кишечнику миші пресинаптичний, синтетичний, постсинаптичний періоди тривають відповідно 9,5; 7,5 та 1 годину); · виду організму |
| постмітотичний (пресинаптичний) (G1 – від англ. gap – проміжок) | Збільшується кількість цитоплазми та органоїдів, а також протікають основні процеси життєдіяльності клітини. | Найдовший період інтерфази (від 50 до 90 % її часу) |
| синтетичний (S – від англ. synthesis – синтез) | Подвоєння (реплікація, редуплікація) ДНК, збільшується кількість РНК, білків-гістонів і інших речовин, що призводить до збільшення розмірів ядра. | Найкоротший період клітинного циклу |
| постсинаптичний (премітотичний) (G2) | Ріст ядра і цитоплазми, накопичення енергії у вигляді молекул АТФ, синтез спеціальних речовини для утворення мітотичного апарату. | 
|
| Мітоз (від грец. мітос – нитка) | Непрямий поділ соматичних еукаріотичних клітин, що складається з ділення ядра (каріокінезу) і ділення цитоплазми (цитокінезу). Він супроводжується морфологічною перебудовою ядер і цитоплазми клітини. Це безперервний процес, у якому кожна стадія непомітно переходить в іншу, але для зручності вивчення виділяють чотири фази: профазу, метафазу, анафазу і телофазу. | |
| Профаза (від грец. про – перед, раніше) | · хромосоми потовщуються внаслідок спіралізації і стають помітні під мікроскопом; · розчиняється ядерна оболонка і хромосоми виходять у цитоплазму; · починає утворюватись веретено поділу (сукупність мікротрубочок, що складаються з білка тубуліну), тобто центріолі клітинного центру розходяться до полюсів клітини | |
| Метафаза (від грец. мета – після, через) | · завершує формуватись веретено проділу; · кожна хромосома прикріплюється центромерою до однієї з мікротрубочок веретена поділу; · усі хромосоми шикуються на „екваторі” клітини, утворюючи метафазну пластинку (у такому стані найкраще вивчати хромосоми) | |
| Анафаза (від грец. ана – знову, поза) | · окремі хроматиди відходять до протилежних полюсів клітини, внаслідок того, що нитки веретена поділу скорочуються і їх розтягують; · біля кожного полюса клітини збирається диплоїдний набір хроматид | |
| Телофаза (від грец. телос – кінець) | · навколо диплоїдного набору хроматид утворюється ядерна оболонка; · хромосоми деспіралізуються; · відбувається цитокінез – поділ цитоплазми |
5. Цитологічні основи статевого розмноження
Статеве розмноження – це виникнення і розвиток нащадків із заплідненої яйцеклітини – зиготи, яка утворюється внаслідок злиття жіночої і чоловічої статевих клітин. У основі розвитку статевих клітин лежить мейоз.
Мейоз – складний непрямий поділ незрілих генеративних клітин, внаслідок якого утворюються статеві клітини – гамети. Це особливий спосіб поділу клітини, у результаті якого відбувається редукція (зменшення) кількості хромосом і перехід клітин з диплоїдного стану у гаплоїдний.
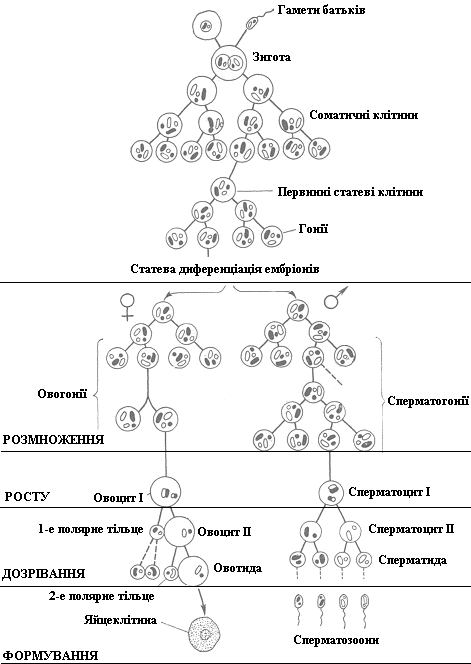
У тварин статеві клітини утворюються з ембріональних, а сам процес утворення називається гаметогенезом.
Гаметогенез (овогенез і сперматогенез) умовно поділяють на періоди розмноження, росту, дозрівання і формування.
Ці два процеси мають свої особливості протікання, але разом з тим вони поділяються на чотири періоди: розмноження; росту; дозрівання; формування.
| Періоди гаметогенезу | Сперматогенез | Овогенез |
| Утворення чоловічих статевих клітин (сперматозоїдів) відбувається в чоловічих статевих залозах – сім’яниках. | Утворення жіночих статевих клітин (яйцеклітин) відбувається у жіночих статевих залозах – яєчниках. | |
| Гамети у тварин (особин жіночої і чоловічої статі) формуються з гоній – первинних диплоїдних статевих клітин, які утворюються в період ембріонального розвитку із зародкових клітин сперматогенного та овогенного епітелію шляхом багаточисленних мітозів. Потім утворення жіночих і чоловічих статевих клітин набуває своїх особливостей. | ||
| Розмноження(мітоз) | Первинні диплоїдні статеві клітини (оогонії чи сперматогонії) діляться шляхом мітозу, перетворюючись в овоцити Ічисперматоцити І | |
| Росту | Утворені клітини (овоцити І чи сперматоцити І) ростуть, збільшуючись у розмірах. | |
| Дозрівання (мейоз) | Сперматоцити І діляться мейозом. Після першого поділу утворюється 2 сперматоцити ІІ, після другого – 4 сперматиди. | Овоцити І діляться мейозом. Після першого поділу утворюється 1 овоцит ІІ та 1-е полярне тільце, після другого – 1 овотида і 2-е полярне тільце, з 1-го полярного тільця утворюються ще два полярних тільця. |
| Формування | Після формування хвостика 4 сперматиди перетворюються на сперматозоїди. | Після формування деяких оболонок овотида перетворюється на яйцеклітину. |
| Паренхіма сім’яника складається з кількох тисяч покручених каналів, які зовні мають сполучнотканинну оболонку. За нею міститься базальна мембрана, а далі – шар диплоїдні клітин сперматогенного епітелію – сперматогонії. Всі періоди сперматогенезу, на відміну від овогенезу, відбуваються після досягнення твариною статевозрілого віку. У диких тварин сперматогенез відбувається у певні періоди року – восени або навесні, а у свійських – цілий рік після досягання статевої зрілості. Інтенсивність сперматогенезу висока. Так, у бугая щодоби утворюється близько 12 млрд сперматозоонів, а це означає, що за 1 с ділиться 32 тис. сперматоцитів І порядку, з яких утворюється близько 130 тис сперматозоонів. | Період розмноження і росту починається і завершується ще в період ембріонального розвитку. Так, наприклад, у майбутньої телиці, а потім і в корови, у 3-місячному віці внутрішньоутробного розвитку утворюється близько 40 тис. овоцитів І порядку. Після досягання організмом статевої зрілості починається період дозрівання та формування, які здійснюється циклічно (у період статевих циклів). У малоплідних тварин кількість яйцеклітин, які дозрівають одночасно, зазвичай становить одну, рідко дві чи декілька, у багатоплідних – зазвичай декілька. | |
| У сперматогенезі: з 1 диплоїдної статевої клітини утворюється 4 сперматозоїди, бо маса сперматозоїда не має важливого значення для розвитку зародка, від постачає лише спадковий матеріал. У овогенезі: з 1 диплоїдної статевої клітини утворюється 1 велика яйцеклітина, яка забезпечує поживними речовинами майбутній зародок, та 4 дрібні полярні тільця, які майже не отримують цитоплазми і потрібні лише для того, щоб звільнити яйцеклітину від зайвих хромосом. |
Мейоз (від грец. мейозіс – зменшення) – складний непрямий поділ незрілих генеративних клітин, внаслідок якого утворюються статеві клітини – гамети, які мають гаплоїдний набір хромосом.
| Період та фаза | Процеси | |
| Редукційний поділ (І) | ||
| Профаза І | Відбуваються процеси, характерні для кожної профази: § хромосоми потовщуються внаслідок спіралізації і стають помітні під мікроскопом; § розчиняється ядерна оболонка і хромосоми виходять у цитоплазму; § починає утворюватись веретено поділу (сукупність мікротрубочок, що складаються з білка тубуліну), тобто центріолі клітинного центру розходяться до полюсів клітини Окрім того відбуваються такі процеси: | 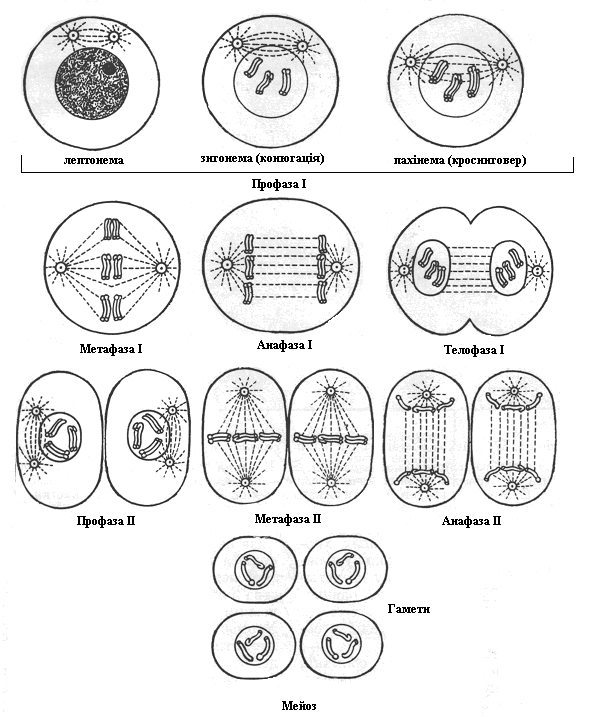
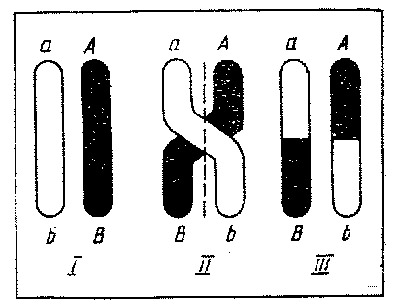 Спрощена схема кросинговеру
Спрощена схема кросинговеру
|
| лептонема (від лат. leptos – тонкий, nema – нитка) | § хромосоми, які складаються з двох хроматид, з’єднаних центромерою, мають вигляд тонких ниток | |
| зигонема (від лат. zygeo – з’єднувати, nema – нитка) | · відбувається кон’югація(від лат. кон’югатіо – сполучення)(процес нагадує застібання змійки) – попарне з’єднання гомологічних хромосом у біваленти (складаються з чотирьох хроматид, які утворюють тетраду); · посилюється спіралізація хромосом, біваленти виглядають як єдине ціле | |
| пахинема (від лат. pachys – товстий, nema – нитка) | · гомологічні хромосоми, з’єднані у біваленти, переплітаються між собою, ще більше спіралізуються, вкорочуються і стають добре помітними у вигляді товстих ниток; · відбуваєтьсякросинговер(crossing over – перехрест) – обмін ділянками між гомологічними хромосомами | |
| диплонема (від. лат. diplos – подвійний, nema – нитка) | § відбувається роз’єднання бівалентів у порядку, зворотному тому, який спостерігався при кон’югації (процес схожий на розстібання “змійки”) – хромосоми відштовхуються одна від одної, тому у цей час під мікроскопом можна побачити перехрести, або хіазми (завдяки схожості хромосом на букву Х), кожна з який відповідає одному акту кросинговеру | |
| діакінез | § закінчується процес спіралізації хромосом і біваленти виглядають як дві дуги, які з’єднані тільки кінцями хромосом | |
| Метафаза І | · до ниток веретена центромерами прикріплюються біваленти (їх удвічі менше диплоїдного набору хромосом) і переміщуються у екваторіальну площину; · центромери гомологічних хромосом розміщуються на протилежних сторонах екваторіальної площини (у метафазі мітозу центромери окремих хромосом розміщуються в екваторіальній площині); · тетради (біваленти) розпадаються на дві діади (дві пари хромосом). | |
| Анафаза І | · до протилежних полюсів розходяться гомологічні хромосоми (діади), хромадити яких завдяки кросинговеру не ідентичні, як на початку мейозу. | |
| Телофаза І | · на протилежних полюсах клітини формуються ядра з гаплоїдним набором хромосом, тобто відбувається редукція кількості хромосом; · зникають нитки веретена, відбувається цитокінез | |
| Інтеркінез | Короткий відпочинок між редукційним поділом, що завершився, і наступним – екваційним. У більшості випадків в інтеркінезі видимих змін в утворених клітинах не спостерігається. У даному випадку хромосоми безпосередньо переходять від телофази І до профази ІІ. Проте у деяких видів у цей час відбувається деспіралізація (розкручування) хромосом. | |
| Екваційний поділ (І) | ||
| Профаза ІІ | · хромосоми спіралізуються, якщо вони деспіралізуються в інтеркінезі; · зникає ядерна оболонка, утворюється веретено поділу | |
| Метафаза ІІ | · хромосоми, які складаються з двох хроматид, прикріплюються центромерами до ниток веретена і вишиковуються в екваторіальній площині | |
| Анафаза ІІ | · центромери діляться, якісно різні хроматиди розходяться і рухаються до протилежних полюсів клітини | |
| Телофаза ІІ | · на протилежних полюсах клітини формуються 4 нових ядра з гаплоїдним набором хроматид, які деспіралізуються; · утворюється ядерна оболонка; · відбувається цитокінез. |
Порівняльна характеристика мітозу та мейозу
| Процеси | Мітоз (непрямий поділ соматичних клітин) | Мейоз (лежить в основі утворення статевих клітин) | |
| перший поділ (редукційний) | другий поділ (екваційний) | ||
| Інтерфаза: синтез ДНК, РНК, білків, подвоєння хромосом, у складі кожної з них з’являється дві хроматиди | Необхідна перед кожним мітозом | Відбувається тільки перед першим поділом | Відсутня |
| Профаза | |||
| Тривалість | Нетривала. | Дуже тривала. | Коротка. |
| Набір хромосом | Диплоїдний набір хромосом, які складаються з двох хроматид. | Диплоїдний набір хромосом (кожна з двох хроматид), з’єднаних у біваленти, кожен з яких складається з чотирьох хроматид. Хроматиди гомологічних хромосом кон’югують і відбувається кросинговер. | Гаплоїдний набір хромосом, які складаються з двох хроматид. |
| Метафаза | |||
| Прикріплення центромер до ниток веретена | До центромери хромосоми | До центромер здвоєних хромосом | До центромери хромосоми |
| Вишиковування хромосом у площині екватора | Хромосоми (кожна з двох хроматид) в диплоїдному наборі | Хромосоми (кожна з двох хроматид) в диплоїдному наборі, з’єднані у бівааленти. | Хромосоми (кожна з двох хроматид) у галоїдному наборі |
| Анафаза | |||
| До протилежних полюсів клітини відходять | Диплоїдні набори хроматид (нових хромосом) | Гаплоїдні набори гомологічних хромосом, кожна з яких складається з двох хроматид | Гаплоїдні набори хроматид (нових хромосом) |
| Телофаза | |||
| Формування клітин у кількості | |||
| Утворюються клітини з набором хромосом | 2n | n | n |
| Біологічне значення | Ø забезпечення сталості генетичної інформації у кількох клітинних поколіннях (сталості каріотипу певного виду) | Ø забезпечення сталості генетичної інформації у кількох поколіннях організмів; Ø підвищення рівня комбінаційної мінливості, що відкриває нові можливості для дії відбору і створення генетично різних форм. Це дозволяє групі тварин існувати в умовах середовища, які постійно змінюються; Ø створення стадійно молодих клітин (гамет) і позбавлення клітини від небезпеки загибелі | |
| Місце у життєвому циклі організмів | – | Ø у споровиків, деяких водоростей (у них більша частина життєвого циклу гаплоїдна) мейоз лежить в основі першого поділу зиготи; Ø у багатоклітинних тварин, голонасінних і покритонасінних мейоз лежить основі утворення статевих клітин; Ø у вищих спорових мейоз лежить в основі утворення спор. |
Оскільки в основі гаметогенезу лежить мейоз, то статеві клітини містять гаплоїдний набір хромосом, який відновлюється до диплоїдного після запліднення.
Запліднення – це процес проникнення (сингамії) сперматозоона в яйцеклітину і злиття (каріогамія) їх ядер. При цьому утворюється зигота з диплоїдним набором хромосом. У результаті каріогамії гомологічні хромосоми, які розходяться у мейозі у попереднього покоління, знову об’єднуються в одному ядрі зиготи.
Незважаючи на величезну кількість сперматозоїдів, що приходяться на одну яйцеклітину, запліднення здійснюється, як правило, лише при участі одного сперматозоїда.
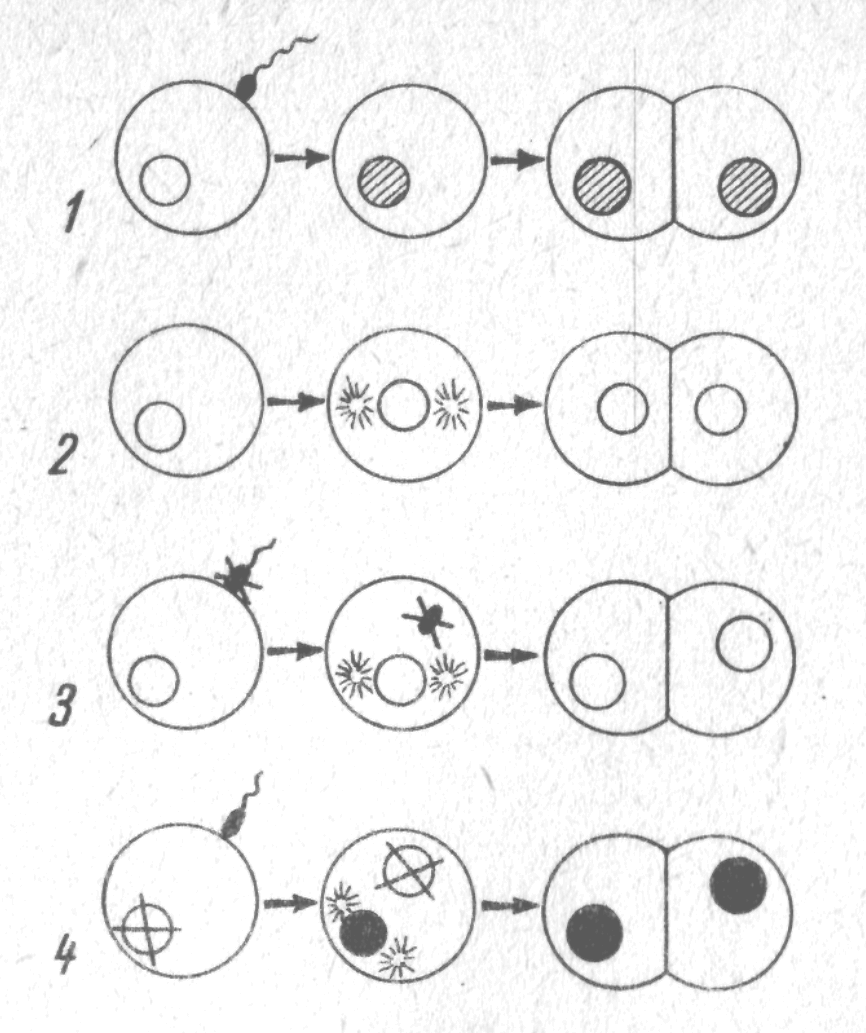
Схема запліднення
Явище, при якому у яйцеклітину проникає лише один сперматозоон, називають моноспермією, декілька – поліспермією.
Поліспермія поширена у безхребетних: молюсків, голкошкірих, комах. Зустрічається у хребетних: риб (в акулових), амфібій, рептилій і птахів. У ссавців у нормі поліспермія зустрічається рідко (1-2%); виключенням є однопрохідні, у яких це явище, можливо, є правилом
У випадку поліспермії, з жіночим пронуклеусом з'єднується тільки один чоловічий пронуклеус, інші елімінуються. Поліспермна каріогамія, тобто злиття декількох пронуклеусів, у нормі не виявлена.
Проникнення в цитоплазму яйцеклітини декількох сперміїв і злиття з її ядром тільки одного з них давно наводили на думку, що даний процес не є чисто механічним. Допускається можливість вибірковості в процесі каріогамії, тобто здатність жіночого пронуклеуса "вибирати" і зливатися з певним чоловічої пронуклеусом.
Рідкісні типи статевого розмноження
| Тип розмноження | Для кого характерний | Схема |
| Партеногенез – розвиток зародка з незаплідненої яйцеклітини. Розрізняють: соматичний чи диплоїдний, і генеративний, чи гаплоїдний. При генеративному (гаплоїдному) зародок розвивається з галоїдної яйцеклітини (у медоносної бджоли трутні розвиваються з незапліднених гаплоїдних яєць шляхом партеногенезу). При соматичному (диплоїдному) яйцеклітина не ділиться, якщо і ділиться, два гаплоїдних ядра, зливаючись разом, відновлюють диплоїдний набір хромосом (автокаріогамія); у такий спосіб у клітинах тканин зародка зберігається диплоїдний набір хромосом. | Нижчі ракоподібні; коловертки; перетинчастокрилі (бджоли, оси) і ін. Відомий у птахів (індички). Партеногенез можна стимулювати штучно, викликаючи активацію незапліднених яєць шляхом впливу різними агентами. Має велике значення для вивчення спадковості, тому що при цьому потомство одержує спадкову інформацію тільки від матері. |
|
| Гіногенез – запліднення не відбувається; розвиток зародка відбувається винятково за рахунок жіночого ядра, як стимулятори розвитку яйцеклітини є сперматозоїди (псевдогамія). | Круглі черви, молінезія, срібний карась. |
|
| Андрогенез – розвиток яйця здійснюється тільки за рахунок чоловічих ядер і материнської цитоплазми. | Шовковичний шовкопряд, паразитична оса (Habrobracon juglandіs). |
|
Date: 2015-09-02; view: 2686; Нарушение авторских прав