Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Свойства нервных центров
|
|
Нервные центры обладают рядом специфических особенностей, обусловленных конструкцией нейронных связей, структурой и свойствами синапсов, характе-
ром и результатом «синаптическои игры» нервных волоконных входов на мембране нейрона-интегратора.
Основные свойства нервных центров -- это одностороннее и замедленное проведение, суммация и трансформация ритма возбуждения, последействие (облегчение), утомление и тонус нервных центров, чувствительность нервных центров к снабжению кислородом и к действию нейротронных веществ, возбуждение и торможение, иррадиация возбуждения, конвергенция и окклюзия, отдача (индукция) и ре-ципрокные отношения, принцип общего конечного пути и принцип обратной связи, принцип доминанты, пластичность нервных центров и их компенсаторные возможности.
Хотя вдоль нервного волокна возбуждение проводится в обе стороны, через синапсы имеет место одностороннее проведение — от пресинаптическои мембраны к постсинаптической, т.е. вдоль рефлекторной дуги от ее афферентного входа к эфферентному концу. Это связано со свойствами синапсов: для химических - - с выработкой медиаторов в пресинаптическои мембране, диффузией их через синапти-ческую щель к хеморецептору постсинаптической мембраны, а для большинства электрических синапсов — с полупроводниковыми свойствами их синапти-ческих мембран.
Замедление проведения возбуждения по рефлекторной дуге обусловлено временной синаптическои задержкой (2—3 мс), включающей время выделения медиатора из пресинаптических везикул, время трансфузии его через синаптическую щель к постсинаптической мембране и время генерации постсинаптичес-кого потенциала (ПСП). При этом истинная синапти-ческая задержка (до начала генерации ПСП) составляет 0,5 мс, а время генерации — еще 1,5-2,5 мс.
Суммация возбуждения, описанная Сеченовым и теоретически рассматриваемая как пространственная и временная, практически представлена пространственно-временной, когда на центральном нейроне конвергируют импульсы, разнесенные пространственно, т.е. поступающие от разных входов и попадающие в сопоставимые с длительностью релаксации ПСП микроинтервалы времени, допускающие их суммацию.
Трансформация ритма возбуждения, т.е. изменение ритма приходящих на входы нейрона импульсных потоков, достигается за счет нескольких механизмов: урежение импульсации связано с более низкой лабильностью нейрона-приемника, обусловленной длительной фазой его следовой гиперполяризации, а учащение, напротив,— с длительной деполяризацией, достигающей критического уровня, что способствует генерации множественных потенциалов действия (как, например, в клетках Ренпюу), а также с включением нейронов в реверберирующие цепочки возбуждения.
Этими же механизмами объясняется и зависимость рефлекторного ответа от силы и длительности раздражения: увеличение этих параметров стимуляции приводит как к включению большего числа нейронов (за счет присоединения к низкопороговым более высокопороговых элементов), так и к суммационно-транс-формационным преобразованиям на синаптических аппаратах центральных нейронов.
Рефлекторное последействие (облегчение) обусловлено (как и при трансформации) длительной следовой критической деполяризацией мембраны нейронов, обеспечивающей кратковременную фасилитацию, и циркуляцией импульсов в «ловушках возбуждения» (Лоренте де Но), создающей условия для длительного последействия (рис. 3.1).
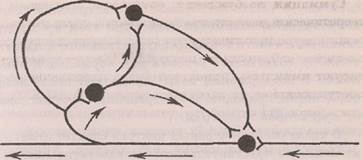
Рис. 3.1. Кольцевые связи нейронов в нервном центре (Лоренте де Но, 1938)
Явлением облегчения объясняется и эффект пост-тетанической потенциации, выражающейся в увеличении ответа (например, передне-корешкового потенциала спинного мозга) после тетанизации рефлекторного входа ритмическими импульсными последовательностями определенной частоты в течение определенного времени (Ллойд).
На нервных центрах можно наблюдать (в отличие от волокон) и эффект утомления, проявляющийся в постепенном снижении и полном прекращении рефлекторного ответа при продолжительном раздражении афферентных входов и являющийся результатом нарушения передачи возбуждения в нейронных синапсах. При этом фазические рефлексы вызывают утомление в нервных центрах довольно быстро, в то время как тонические могут длительно протекать, не сопровождаясь развитием утомления. Это создает возможности для многочасового поддержания позного мышечного тонуса, что, в свою очередь, по путям обратной афферентации обусловливает тонус нервных центров, обеспечивающих постоянную (но разной интенсивности в зависимости от состояния на периферии) импульсацию к соответствующим периферическим
системам, а также постоянное межцентральное (меж-нейрональное) взаимодействие. При этом все проявления мышечного тонуса (контрактильного и пластического), создающего условия для нормальной локо-моции и поддержания позы, пропадают после деаф-ферентации, приводящей к устранению обратной связи от эффекторов к центрам.
В числе свойств нервных центров следует отметить их высокую чувствительность к недостатку кислорода (которая понижается при гипотермии, приводящей к снижению уровня метаболизма) и к ряду нейротроп-ных веществ: нервным ядам, наркотикам, алкоголю, ганглиоблокаторам, антидепрессантам, психостимуляторам, транквилизаторам.
Интегративные функции нервных центров обеспечиваются специфическими закономерностями во взаимодействии возбудительного и тормозного процессов, причем торможение часто играет ведущую роль в достижении координированной деятельности центральной нервной системы. Центральное торможение (впервые описано И. М. Сеченовым) представляет собой особый нервный процесс, вызываемый возбуждением и проявляющийся в подавлении другого возбуждения.
Различают первичное торможение, являющееся результатом активации особых тормозящих структур, действующих на тормозимую клетку и вызывающих в ней торможение как первичный процесс без предварительного возбуждения, и вторичное торможение, возникающее в клетке без действия на нее специфических тормозящих структур, а как следствие ее возбуждения, т.е. вторично. К первичному торможению относятся постсинаптическое торможение и пресинаптичес-кое (разновидностью постсинаптического являются возвратное и латеральное торможение). К вторичному — пессимальное и торможение вслед за возбуждением.
Наличие в нервной системе тормозящих синапсов было показано Ллойдом и Реншоу. Основой синапти-ческого торможения является гиперполяризация по-стсинаптической мембраны тормозимого нейрона, представляющая собой постсинаптический потенциал (ТПСП). В качестве медиатора постсинаптического торможения могут выступать фактор I (Флори), гам-ма-аминомасляная кислота (ГАМК), норадреналин, серотонин, ацетилхолин, аминокислоты и пептиды, приводящие к изменению хлорной и калиевой проницаемости. Характер постсинаптического процесса определяется взаимодействием медиатора с хеморе-цепторными структурами постсинаптической мембраны, в результате чего возникает гиперполяризация (ТПСП) либо деполяризация (ВПСП). Описаны специфические тормозные нейроны, которые, возбуждаясь, выделяют в своих пресинаптических окончаниях тормозной медиатор. К таким нейронам относятся спинномозговые клетки Реншоу, синаптирующие на возбуждающих их мотонейронах и вызывающие эффект возвратного торможения; подобные интернейроны описаны и в других отделах мозга. В зависимости от характера хеморецептора один и тот же медиатор может вызвать у одних нейронов генерацию ВПСП, а у других — ТПСП. Поскольку на постсинаптической мембране одного и того же нейрона взаимодействуют от многих синаптических входов множественные ВПСП и ТПСП, обеспечивая интегративную деятельность нейрона, постсинаптическое торможение получило название координационного. Постсинаптическое торможение развивается в основном в аксодендрит-ных синапсах.
Другой вид первичного торможения -- пресинап-тическое торможение — развивается за счет деполяризации пресинаптических аксонных терминален (по-
добно катодической депрессии Вериго) из особых тормозных синапсов возбужденного интернейрона. В результате возникает частичная или полная блокада проведения возбуждения к нервным окончаниям, си-наптирующим на данном (тормозимом) нейроне. Поскольку пресинаптическое торможение не участвует, подобно постсинаптическому, в «синаптической игре», а лишь блокирует синаптические входы, то оно называется фильтрационным. Пресинаптическое торможение связано с аксо-аксональными синапсами.
Вторичное пессимальное торможение развивается в возбуждающих синапсах в результате сильной и длительной деполяризации синаптической мембраны приходящими сюда высокочастотными импульсными потоками (подобно пессимуму Введенского в нервно-мышечном синапсе либо катодической депрессии Вериго), что приводит к аккомодационным изменениям и повышению порога возбуждения в постсинаптичес-кой мембране, делающим ее инактивной.
Торможение вслед за возбуждением развивается в нейроне в результате возникновения поствозбудительной следовой гиперполяризации его мембраны.
ТПСП, как и ВПСП, представляет собой локальный процесс. Однако ВПСП, в отличие от ТПСП, достигая критической величины деполяризации постси-наптической мембраны, превращается в распространяющийся вдоль нервного волокна потенциал действия, что приводит к новым синаптическим процессам — генерации очередного ВПСП или ТПСП.
Распространение возбуждения в центральной нервной системе, названное иррадиацией (Гольтц, Шер-рингтон), достигается за счет многочисленных волоконных ветвлений и при активном участии ретикулярной формации, а на уровне спинного мозга — желати-нозной субстанции Роландо. Ограничение иррадиации
осуществляется благодаря механизму возвратного торможения с помощью специальных тормозных интернейронов (на уровне спинного мозга эту роль играют клетки Реншоу).
Важным механизмом координации рефлекторных процессов является конвергенция (Шеррингтон), которая достигается схождением к одним и тем же промежуточным или эффекторным нейронам различных афферентов от разных участков общего рецептивного поля (в спинном и продолговатом мозгу) или даже от разных рецептивных полей (в высших отделах головного мозга). При этом могут иметь место два эффекта: 1) окклюзия (закупорка) в случае конвергенции на нейроне импульсных потоков, каждый из которых способен обеспечить пороговое или сверхпороговое раздражение адресного нейрона (по закону «все или ничего» в данном случае суммации не происходит), и тогда суммарный эффект будет меньше арифметической суммы двух эффектов; 2) облегчение в случае конвергенции на адресном нейроне таких импульсных последовательностей, каждая из которых не может создать порогового воздействия, и тогда эти два или несколько эффектов суммируются.
Шеррингтоном были также описаны возбудительно-тормозные отношения в спинном мозгу (а Введенским — в двигательной коре), приводящие к координации флексорно-экстензорных взаимодействий и названные реципрокными (взаимосочетанными, или сопряженными). В основе таких реципрокных рефлексов лежит постсинаптическое торможение, возникающее (через вставочный тормозной нейрон) в мотонейроне, иннервирующем мышцу-экстензор при рефлекторном возбуждении другого мотонейрона, иннервирующего соответствующую антагонистическую мышцу-флексор. Одновременно на контралатеральной стороне возникают
противоположные реакции — возбуждение экстензора и торможение флексора. Это перекрестный разгибатель-ный рефлекс. Такие взаимотормозящие влияния рефлексов, основанные на реципрокном торможении, обусловливают состояние одновременной отрицательной индукции. А последовательные смены возбуждения и торможения — состояние последовательной положительной (возбуждение вслед за торможением) и последовательной отрицательной (торможение вслед за возбуждением) индукции (И. М. Сеченов, Шеррингтон). Эта смена возбуждения и торможения в нервных центрах лежит в основе феномена «отдачи» (Шеррингтон) и ритмических цепных рефлексов, обеспечивающих акты шагания, чесания и др.
При этом, поскольку один и тот же нейрон (эффек-торный либо промежуточный) может входить в разные «вероятностно-статистические ансамбли» (А. Б. Коган) и участвовать в осуществлении различных функций, возникает «борьба за центры», образующие общий конечный путь (Шеррингтон). А поскольку эф-фекторных нейронов примерно в 5 раз меньше, чем рецепторных, то это схождение путей к общему конечному получило название «принципа воронки» (Шеррингтон). Рефлексы, имеющие общий конечный путь, подразделяются на алиированные (союзные), взаимно подкрепляющие и усиливающие друг друга, и антагонистические, конкурентные и тормозящие друг друга.
Адекватная динамика возбудительно-тормозных состояний центров обеспечивается поступлением вторичных афферентных импульсов по путям обратных аффе-рентаций от рецепторов работающих органов, осуществляющих обратную связь (И. М. Сеченов, Шеррингтон, Бернштейн, П. К. Анохин), корригирующую через акцептор действия программу действия (П. К. Анохин).
Очень важным рабочим принципом нервных центров является принцип доминанты (А. А. Ухтомский). Доминантный очаг характеризуется, по А. А. Ухтомскому, повышенной возбудимостью, стойкостью возбуждения, способностью к суммированию возбуждений, инерцией, способностью как бы «притягивать» возбуждения от других нервных центров, усиливаясь за их счет и подавляя их реакцию индукционным торможением. В. С. Русиновым было показано более раннее срабатывание искусственно созданного с помощью поляризации доминантного очага по сравнению с другими корковыми проекционными зонами, куда собственно адресовался сенсорный сигнал; это может быть объяснено с позиций теории конвергентного замыкания (П. К. Анохин).
Нервные центры обладают довольно высокой пластичностью (Бете), обеспечивающей приспособляемость нервной системы и ее компенсаторные возможности в случае каких-либо возникающих нарушений функций (И. П. Павлов, П. К. Анохин, Э. А. Асра-тян, А. Б. Коган).
Date: 2016-11-17; view: 532; Нарушение авторских прав