Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Морфофункциональная организация
|
|
Спинной мозг, являясь самым каудальным отделом центральной нервной системы, осуществляет две основные функции — рефлекторную и проводниковую. Импульсы, приходящие от экстерорецепторов кожи, проприо- и висцерорецепторов, обеспечивают разнообразие двигательных и вегетативных рефлекторных реакций, в характере протекания которых не последнюю роль играют межнейронные отношения в спинном мозгу и межцентральные отношения спинного и головного мозга, что обеспечивается внутриспиналь-ными межнейронными связями и восходящими и нисходящими проводящими путями спинного мозга.
Различные представители позвоночных обладают спинным мозгом неодинаковой длины. Так, у большинства рыб, хвостатых амфибий, рептилий и птиц он заполняет весь позвоночный канал, в то время как у бесхвостых амфибий и млекопитающих -- только часть этого канала. Самый короткий спинной мозг обнаружен у низших животных. Так, например, у южноамериканской жабы (Р1ра) конечный конус спинного мозга находится на уровне третьего позвонка, а у высокодифференцированной костистой рыбы (Мо1а) спинной мозг вообще не выходит за пределы черепа, и весь позвоночный канал заполнен лишь нервными волокнами.
Спинной мозг имеет сегментарное строение. От каждого сегмента отходят две пары корешков — вентраль-
ные и дорсальные, которые, соединяясь, образуют периферические спинномозговые нервы. В опытах с перерезками спинальных корешков, раздражением их и отведением от них электрической активности было показано, что задние корешки являются афферентными, чувствительными, центростремительными, а передние — эффекторными, двигательными (либо секреторными), центробежными -- это закон распределения корешков Белла—Мажанди. Перерезка задних корешков приводит не только к потере чувствительности, но и к нарушениям органов движения, которые становятся плохо координированными -- атаксическими. Спинномозговая атаксия возникает и у человека при поражениях восходящих путей спинного мозга. Такие расстройства координации движений наступают вследствие неправильного распределения мышечного тонуса в результате потери обратной связи от проприоре-цепторов и экстерорецепторов кожи во время осуществления двигательного акта.
Шеррингтоном в опытах с перерезками спинальных корешков было показано, что каждый сегмент спинного мозга иннервирует три метамера тела и получает афферентацию от трех метамеров, а каждый метамер иннервируется из трех сегментов спинного мозга и передает сигналы в три спинномозговых сегмента.
Соответственно распределению и функциям корешков на поперечном сечении спинного мозга различаются задние и передние рога центрального серого вещества, имеющего Н-образную форму. В задние рога входят волокна задних корешков (тела нейронов которых локализованы в спинальных межпозвоночных ганглиях), здесь распределены тела промежуточных нейронов (интернейронов) и находятся клетки жела-тинозной субстанции Роландо; в передних рогах ло-
|
| Кортикоспинальный тракт |
| Руброспияальный тракт |
| Латеральный р цти к улосп ин ал ьн ый тракт |
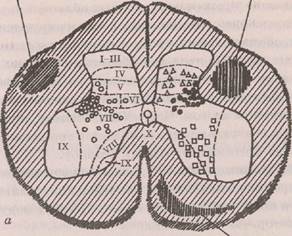
кализованы двигательные нейроны (мотонейроны), волокна которых покидают спинной мозг в составе передних корешков. Между дорсальными и вентральными рогами лежит промежуточная зона. Нервные клетки распределены и диффузно, и ядерно: медиальные и латеральные ядра — в переднем роге, ин-термедиомедиальное и интермедиолатеральное ядра — в промежуточной зоне, кларков столб и собственное ядро — в заднем роге. Интернейроны имеют небольшие размеры сомы, короткие дендриты и аксоны различной длины. Мотонейроны обладают крупной сомой, длинными ветвящимися дендритами, проникающими за пределы серого вещества, и длинными толстыми аксонами, иннервирующими мышцы. В боковых рогах спинного мозга локализованы вегетативные нейроны, представляющие собой инстанцию исполнительного механизма в регуляции вегетативных функций. В сером веществе спинного мозга интер- и мотонейроны образуют функциональные объединения, отвечающие за определенные специфические функции,— это функциональные единицы центральной нервной системы. Рекседом предложено разделение на XI пластинок (или слоев) серого вещества спинного мозга по анатомо-функциональным особенностям составляющих их нейронных элементов (рис. 4.1).
1-ГУ пластинки, образующие «головку дорсального рога», являются первичной сенсорной областью серого вещества для проекции большей части экстеро-цептивных афферентных волокон от туловища и конечностей. Из этой области идут тракты, передающие экстероцептивные сигналы к высшим центрам.
У-У1 пластинки образуют «шейку дорсального рога», где оканчиваются волокна от сенсомоторных областей коры и волокна проприоцептивной чувствительности туловища и конечностей; кроме того, в
Вестибулоспинальный тракт
 Латеральный ретикулоспиналыгый тракт (
Латеральный ретикулоспиналыгый тракт (
Медиальный ретикулоспиналышй тракт
Рис. 4.1. Схема разделения серого вещества спинного мозга у поясничного сегмента кошки (Рексед, 1964): а — расположение и области окончаний: слева — руброспинального тракта (светлые кружки); справа — вестибулоспинального (квадраты) а кортикоспинального трактов (треугольники и черные кружки); б — расположение окончаний медуллярного (светлые кружки) и мостового (треугольники) ретикулоспаналь-ных трактов (Шмидт, Тевс, 1983)
медиальную часть V (и IV) пластинки проецируются нисходящие волокна от сенсорных областей коры. По предположению Рекседа, здесь интегрируются рефлексы, обеспечивающие точную регуляцию движений.
Пластинка VII является областью обширных про-прио- и висцероспинальных связей, а также афферентных и эфферентных связей спинного мозга с мозжечком и средним мозгом. Часть VII пластинки, локализованная в вентральном роге, является тормозной областью, где преобладают клетки Реншоу.
Нейроны пластинки VIII, характеризующейся буль-бо- и проприоспинальными (особенно контралатераль-ными и комиссуральными) связями, могут модулировать моторную активность.
Пластинка IX является первичной моторной областью серого вещества и содержит мотонейроны спинного мозга, объединенные в функциональные группы (пулы) и представляющие собой общий конечный путь рефлексов спинного мозга.
Клетки нейроглии спинного мозга на значительном протяжении покрывают поверхность нейронов, причем отростки глиальной клетки направлены, с одной стороны, к телам нейронов, а с другой, часто контактируют с кровеносными капиллярами, являясь, таким образом, посредниками между нервными элементами и источниками их питания (Г. И. Поляков).
Спинной мозг по восходящим путям передает сигналы в надсегментарные уровни головного мозга, а но нисходящим получает оттуда команды к действию. Восходящие пути передают импульсы от проприоцеп-торов по волокнам спинобульбарных пучков Голля и Бурдаха (задние столбы белого вещества) и спиномоз-жечковых путей Говерса и Флексига (боковые столбы), от болевых и температурных рецепторов по латеральному спиноталамическому тракту, от тактильных
рецепторов по вентральному спиноталамическому пути и частично по пучкам Голля и Бурдаха.
Нисходящие пути проходят в составе кортикоспи-нальных, или пирамидных, трактов (передние и боковые столбы) и экстракортикоспинальных, или экстрапирамидных: ретикулоспинального, руброспи-нального, тектоспинального, вестибулоспинального, оливоспинального пучков.
Date: 2016-11-17; view: 652; Нарушение авторских прав