Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Нейропсихологииеские методы
|
|
К методам этой группы относятся, как правило, комплексные методы объективной оценки свойств ЦНС, связанных с психическими актами. К ним можно отнести ощущения, восприятие, внимание, память, мышление, произвольные двигательные реакции.
Поэтому эти методы применяются в равной мере как физиологами, так и психологами — все зависит от поставленной задачи. Объективизация результатов зачастую позволяет выделять отдельные компоненты нервно-психических актов, разделяя в них физиологические и психические звенья.
Рассмотрим это на примере методики измерения времени двигательной реакции (ВДР). В самом общем виде по степени сложности ВДР можно разделить на три класса: простая реакция, реакция различения и
реакция выбора. Простой реакцией называют такую, которая осуществляется при предъявлении одного заранее известного сигнала и получении одного определенного двигательного ответа. Например, в ответ на световой сигнал (вспышка) испытуемый должен как можно быстрее нажать на кнопку. Реакция различения отличается от первой тем, что предъявляемых сигналов бывает несколько, а ответ — только на один из них. Реакция выбора — наиболее сложная. В этом случае испытуемому предъявляется одновременно несколько сигналов, и на каждый из них он должен отвечать определенным действием. Показано, что ВДР заметно увеличивается от степени сложности задания. Так, время простой зрительно-моторной реакции составляет 150—220 мс, а реакция выбора — 1 000 мс и более. Как показали исследования, эта прибавка связана в основном с этапом принятия решения (психическое звено) при практически неизменном времени на обеспечение фотохимических реакций в сетчатке, развитие и распространение возбуждения в нервных проводниках и мышцах.
Среди методов оценки ощущений можно выделить методы изучения адаптации и сенсибилизации зрительного, слухового, тактильного анализаторов, измерение абсолютного и дифференциального порогов чувствительности.
Для анализа восприятия применяются методы кратковременных экспозиций и определения объема восприятия, изучения бинокулярного зрения и бина-уралъного слуха, стереоскопические эффекты. В целях изучения внимания и памяти разработаны методы, с помощью которых возможны оценка закономерностей распределения и переключения внимания, исследование кратковременной памяти, процессов обучения, воспроизведения и узнавания. К этой группе
можно отнести также методы изучения условнореф-лекторной деятельности.
2. Основы физиологии нейрона, глии, синапса,
Физиология нейрона.
Основными элементами нейронной системы являются нервные клетки. Подтверждение клеточной теории строения нервной системы было получено с помощью электронной микроскопии, показавшей, что мембрана нервной клетки напоминает основную мембрану других клеток. Она представляется сплошной на всем протяжении поверхности нервной клетки и отделяет ее от других клеток. Каждая нервная клетка является анатомической, генетической и метаболической единицей так же, как и клетки других тканей организма. Понятие, что одиночная нервная клетка служит, основной функциональной единицей, сменилось представлением о том, что такой функциональной единицей является ансамбль тесно связанных друг с другом нейронов. Нервная система состоит из популяций таких единиц, которые организованы в функциональные объединения разной степени сложности. В нервной системе человека содержится около 100 млрд нервных клеток. Поскольку каждая нервная клетка функционально связана с тысячами других нейронов, то количество возможных вариантов таких связей близко к бесконечности. Нервную клетку следует рассматривать как один из уровней организации нерв-: ной системы, связующих молекулярный, синаптичее-кие, субклеточные уровни с надклеточными уровнями локальных нейронных сетей, нервных центров и функциональных систем мозга, организующих поведение.
Нервные клетки выполняют ряд общих неспецифических функций, направленных на поддержание собственных процессов организации. Это обмен веществами с окружающей средой, образование и расходование энергии, синтез белков и др. Кроме того, нервные клетки выполняют свойственные только им специфические функции по восприятию, переработке и хранению информации. Нейроны способны воспринимать информацию, перерабатывать (кодировать) ее, быстро передавать информацию по конкретным путям, организовывать взаимодействие с другими нервными клетками, хранить информацию и генерировать ее. Для выполнения этих функций нейроны имеют полярную организацию с разделением входов и выходов и содержат ряд структурно-функциональных частей.
Тело нейрона, которое связано с отростками, является центральной частью нейрона и обеспечивает питанием остальные части клетки. Тело покрыто слоистой мембраной, которая представляет собой два слоя липидов с противоположной ориентацией, образующих матрикс, в который заключены белки. Часть мембранных белков является гликопротеинами с полиса-харидными цепочками, выступающими над наружной поверхностью мембраны. Они вместе с углеводами образуют гликокаликс — тонкий слой на поверхности клеточной мембраны, который заполняет межклеточные щели и способствует созданию связей между нейронами, распознаванию клеток, регуляции диффузии через мембрану, обмену с внешней средой. Тело нейрона имеет ядро или ядра, содержащие генетический материал (рис. 2.1),
Ядро регулирует синтез белков во всей клетке и контролирует дифференцирование молодых нервных клеток. При усилении активности нейрона увеличивается площадь ядра и активизируются ядерно-
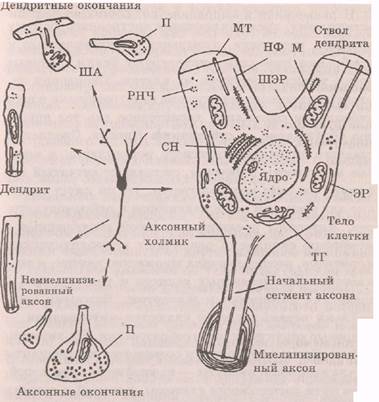
Рис 2.1. Схемы структурно-функциональных частей нейрона. Нейрон, показанный в центре, окружен схемами, иллюстрирующими ультраструктуру его частей: ЭР — эндоплазмати-ческий ретикулюм; ШЭР — шероховатый эндоплазматичес-кий ретикулюм; ТГ ~ тельце Гольджи; СН — субстанция Ниссля; МТ — микротрубочка; НФ — пейрофиламент; М — митохондрия; РНЧ -- рибонуклеопротеиновые частицы; ША — шипиковый аппарат; П — пузырьки (Шеперд, 1987)
плазменные отношения. В цитоплазме тела нейрона содержится большое количество рибосом. Одни рибосомы располагаются свободно в цитоплазме по одной или образуют скопления -- «розетки», где синтезируются белки, которые остаются в клетке. Другие
рибосомы прикрепляются к эидонлазматическому
ретикулюму, представляющему внутреннюю систему мембран, канальцев, пузырьков. Прикрепленные к мембранам рибосомы синтезируют белки, которые потом транспортируются из клетки. Скопления эн-доплазматического ретикулюма со встроенными в него рибосомами составляют характерное для тел нейронов образование — субстанцию Ниссля. Скопления гладкого эндонлазматического ретикулюма, в которые не встроены рибосомы, составляют сетчатый аи-парат Голъджи; предполагается, что он имеет значение для секреции неиромедиаторов и нейромодулято-ров. Лизосомы представляют собой заключенные в мембраны скопления различных гидролитических ферментов, расщепляющих множество внутри- и вне-клеточ ноле каля зова иных веществ и участвующих в процессах фагоцитоза и экзоцитоза. Важными орга-неллами нервных клеток являются митохондрии — основные структуры энергообразования. На внутренней мембране митохондрии содержатся все ферменты цикла лимонной кислоты — важнейшего звена аэробного пути расщепления глюкозы, который в десятки раз эффективней анаэробного пути. Ферменты цепи переноса электронов создают энергию, которая идет на образование АТФ и АДФ. Важной особенностью анергетического обмена нервных клеток является отсутствие собственных углеводов в форме гликогена. Нейроны позвоночных используют глюкозу, беспозвоночных — трегалозу. Высокий уровень энерготрат нервных клеток и отсутствие собственных запасов углеводов делают их особо чувствительными к нарушению поступления крови, в которой содержится глюкоза и кислород, необходимые для аэробного энергообразования на митохондриях. В нервных клетках содержатся также микротрубочкк, нейрофиламенты и
микрофиламенты, различающиеся диаметром. Мжк-ротрубочки (диаметр 300 нм) идут от тела нервной клетки в аксон и дендриты и представляют собой внутриклеточную транспортную систему. Нейрофиламенты (диаметр 100 нм) встречаются только в нервных клетках, особенно в крупных аксонах» и тоже составляют часть ее транспортной системы. Микрофиламен-ты (диаметр 50 нм) хорошо выражены в растущих отростках нервных клеток, они участвуют в некоторых видах межнейронных соединений.
Дендриты представляют собой древовидно»ветвящиеся отростки нейрона, его главное рецептивное поле, обеспечивающее сбор информации, которая поступает через синапсы от других нейронов или прямо из среды. При удалении от тела происходит ветвление дендритов: число дендритных ветвей увеличивается, а диаметр их сужается. На поверхности дендритов многих нейронов (пирамидные нейроны коры„ клетки Пуркинье мозжечка и др.) имеются шипики. Шипиковый аппарат является составной частью системы канальцев дендрита: в дендритах содержатся микротрубочки, нейрофиламенты, сетчатый аппарат Гольджи и рибосомы. Функциональное созревание и начало активной деятельности нервных клеток совпадает с появлением шипиков; продолжи тельное прекращение поступления информации к нейрону ведет к рассасыванию шшшков. Наличие шипиков увеличивает воспринимающую поверхность дендритов; так, площадь дендритов клеток Пуркжнье мозжечка около 250 000 мкм2. Мембрана дендритов по своим свойствам отличается от мембраны других участков нервной клетки и не способна к быстрому и надежному проведению возбуждения.
Аксон представляет собой одиночный,, обычно длинный выходной отросток нейрона, служащий для быс-
трого проведения возбуждения. (В структуру аксона входят начальный сегмент, аксональное волокно и телодендрий.) Аксональное волокно отличается постоянством диаметра по всей длине. В конце он может ветвиться на большое (до 1000) количество веточек. Аксоплазма содержит множество микротрубочек и нейрофиламентов, с помощью которых осуществляется аксональныи транспорт химических веществ от тела к окончаниям (ортоградный) и от окончаний к телу нейрона (ретроградный). Существует быстрый аксональныи транспорт со скоростью сотен миллиметров в сутки и медленный транспорт со скоростью несколько миллиметров в сутки. По аксону транспортируются вещества, необходимые для синаптической передачи, пептиды, продукты нейросекреции. В зависимости от скорости проведения возбуждения различают несколько типов аксонов, отличающихся диаметром, наличием или отсутствием миелиновой оболочки и другими характеристиками (табл. 2.1).
Таблица 2.1 Характеристика разных типов аксонов
| Тип аксона | Скорость, м/с | Диаметр, мкм | Наличие миелиновой оболочки |
| А а | 420-70 | 22-12 | Есть |
| Ар | 70-40 | 12-8 | » |
| Ау | 40-15 | 8-4 | » |
| А8 | 15-6 | 4-1 | » |
| В | 18-3 | 3-1 | » |
| С | 3-0,5 | 2-0,5 | Нет |
Начальный сегмент аксона нейронов является тригерной зоной -- местом первоначальной генерации возбуждения. Этот участок нервной клетки начинается от аксонного холмика и, воронкообразно
сужаясь, переходит в начальный участок аксона, не покрытый миелиновой оболочкой. Поскольку этот участок мембраны нейрона является наиболее возбудимым, то здесь обычно первоначально и возникает возбуждение, которое затем распространяется по аксону и телу нейрона. Таких запускающих возбуждение участков может быть несколько. Начальный сегмент аксона имеет важное значение для интегратив-ной деятельности нервной клетки. Телодендрий представляет собой часть нервной клетки, которая осуществляет соединение с другими нейронами путем синаптических контактов. Это конечные разветвления — терминали аксона, которые не покрыты миелиновой оболочкой и заканчиваются утолщениями различной формы (булавы, кольца/пуговки, чаши и др.), которые входят составной частью в синапс. В утолщениях локализовано значительное количество пузырьков, расположенных свободно или встроенных в пресинаптические мембраны. Поскольку терминали аксона очень тонкие и не покрыты миелином, то скорость возбуждения в них значительно меньше, чем в аксонах.
Взаимодействие частей нервных клеток обеспечивает реализацию их функций с помощью химических и электрических процессов. Химические процессы в нервных клетках отличаются высокой интенсивностью, сложностью и многообразием. Наряду с уже отмеченными особенностями энергетического обмена, в нервных клетках происходит синтез белков (в том числе специфических) широкого спектра, функционально активных пептидов, медиаторов и модуляторов синаптических процессов, продуктов нейросекреции. Электрические процессы имеют важнейшее значение для информационной деятельности нервных клеток и должны быть рассмотрены отдельно.
2.2. Электрические процессы в нейронах
Электрические процессы в нервных клетках включают в себя наличие постоянного потенциала покоя и медленных и быстрых изменений этого потенциала при возбуждении. Потенциал покоя является мембранным потенциалом нервной клетки и обусловлен неравномерным распределением электролитов по обе стороны клеточной мембраны. Внутри нервной клетки содержится большое количество органических анионов и катионов; в наружной среде катионов К+ при* мерно в 40 раз меньше, но высока концентрация катионов Ка+, анионов С1~. Крупные органические анионы не проникают через мембрану, а ионы К+, легко проникающие через мембрану, по закону диффузии перемещаются из области более высокой концентрации наружу. Это приводит к избытку положительных зарядов на наружной поверхности и преобладанию отрицательных зарядов на внутренней поверхности мембраны. Внутренняя поверхность мембраны заряжается отрицательно по отношению к наружной, при этом возникает электрическая сила, обеспечивающая обратное движение части ионов К* внутрь клетки, и устанавливается определенное равновесие, при котором суммарный поток ионов через мембрану будет равен нулю. Разность потенциалов между двумя сторонами мембраны при таком равновесии определяет величину мембранного потенциала. Наряду с потоками ионов Кь, являющихся основными факторами мембранного потенциала, через мембрану нервной клетки в значительно меньшем количестве движутся ионы Ма+, Са++, С1. Они проходят через двойной ли-пидный слой мембраны по своим специальным для каждого вида ионов каналам, открывание и закрывание которых связано с изменением величины мембранного потенциала.
Влияние разницы концентраций и проницаемости основных ионов, участвующих в образовании мембранного потенциала, выражено в уравнении постоянного поля:
ЕМ = ………………………………….
Для создания разницы ионных концентраций и восполнения потерь ионов в мембране нервной клетки действует система мембранного насоса, осуществляющего активный транспорт ионов против градиента концентрации и использующего для этого энергию нейронного метаболизма. Наиболее существен натрий-калиевый насос, возвращающий К+ внутрь клетки и выводящий из нее Ма4. На внутренней стороне мембраны Ма+ соединяется с молекулой переносчика; образованный комплекс ион-переносчик проходит через мембрану; на наружной поверхности комплекс распадается, высвобождая ион N3^ и соединяясь с ионом К+, транспортирует его внутрь. Источником энергии для работы насоса служит расщепление АТФ ферментом АТФ-азой, выполняющим функцию переносчика.
Поскольку соотношение количества переносимых насосом Ма+ и К4 неодинаково, то насос не только поддерживает разницу ионных концентраций по обе стороны мембраны, но и участвует в формировании потенциала покоя, является электрогенным. Таким образом, мембранный потенциал создается в результате работы пассивных и активных механизмов, соотношение которых у разных нейронов неодинаково. Поэтому у различных нейронов величина мембранного потенциала колеблется от -80 до -40 мв, она в значительной степени зависит от особенностей его дея-
тельности и функционального состояния. При уменьшении величины мембранного потенциала покоя (деполяризации) возбудимость возрастает, при увеличении мембранного потенциала (гиперполяризации) возбудимость снижается. Возбуждение нервной клетки связано с развитием потенциала действия. Потенциал действия, или нервный импульс, представляет собой кратковременное, длящееся миллисекунды изменение мембранного потенциала, при котором уменьшается его величина, доходит до нуля и затем потенциал меняет знак. В момент пика потенциала действия мембрана становится заряженной внутри не отрицательно, а положительно (4—50 мв); амплитуда потенциала действия составляет 110—130 мв.
Перезарядка мембраны при возбуждении происходит из-за быстрого и значительного повышения мембранной проницаемости для N3% вследствие чего большое количество ионов Ыа+ проникает с наружной на внутреннюю сторону мембраны и создает здесь избыток положительных зарядов (рис. 2.2).

Рис. 2.2. Происхождение потенциала действия: а — связи между деполяризацией мембраны, увеличением натриевой проницаемости и входящим током Nа*; б — кривые изменения, ионных проницаемостей в процессе формирования потенциала действия (Щеперд, 1987)
Восходящая фаза потенциала действия обусловлена избирательным повышением проницаемости мембраны для Ма+. Раскрытие натриевых каналов связано с уменьшением мембранного потенциала и происходит со все возрастающей интенсивностью — лавинообразно, так как переход Ма+ на внутреннюю поверхность усиливает деполяризацию и приводит к раскрытию новых натриевых каналов. Нисходящая фаза потенциала действия связана с инактивацией натриевых каналов и повышением проницаемости для К+, так как калиевые каналы раскрываются позже натриевых.
Усиленный поток 1С наружу приводит к восстановлению мембранного потенциала до величины потенциала покоя. В телах многих нейронов потенциал действия связан и с входящим током Са++, отличающимся большей продолжительностью. Вход Са++ внутрь клетки во время потенциала действия является эффективным механизмом повышения внутриклеточной концентрации свободного Са++, который запускает или участвует в работе многих метаболических процессов. Во время возбуждения значительно усиливается работа натрий-калиевого насоса, активируемая повышением концентрации Ка+ на внутренней поверхности мембраны. Его деятельность способствует восстановлению потенциала покоя. Потенциал действия обладает порогом, при котором деполяризация достигает критического уровня и раскрываются все натриевые каналы мембраны. При под-пороговых воздействиях раскрывается лишь часть натриевых каналов, перезарядка мембраны не происходит, возникает местное возбуждение. Вследствие того, что при потенциале действия раскрываются все натриевые каналы, его амплитуда постоянна и не зависит от силы раздражения; с этим связана и невосприимчивость к новому раздражению. Потенциа-
лы действия способны быстро и надежно распространяться по мембране тела и аксона нервной клетки. Способность к распространению возбуждения связана с тем, что во время потенциала действия происходит изменение знака заряда в возбужденном участке мембраны. Между ним и невозбужденными соседними участками мембраны возникают локальные электрические токи, под действием которых происходит деполяризация новых соседних участков, что приводит к формированию в них потенциала действия. Далее развиваются локальные токи между новым участком, охваченным возбуждением, и следующими невозбужденными участками; и так возбуждение активно распространяется вдоль всей не-миелинизированной мембраны. Чем больше диаметр волокна, тем скорость распространения возбуждения выше (см. табл. 2.1).
У позвоночных большинство аксонов покрыто ми-елиновой оболочкой, периодически прерывающейся на перехватах Ранвье. В перехватах существует высокая плотность потенциалзависимых натриевых каналов (12 000 на 1 мм2), здесь генерируется потенциал действия, а на участках между перехватами возможно электротоническое формирование локальных токов, вызывающих потенциал действия лишь на следующем перехвате. Благодаря этому происходит скачкообразное (сальтаторное) распространение потенциала действия со значительно большей скоростью, чем по немиелинизированной мембране. Разновидность активного проведения возбуждения выявлена и на определенных участках дендритов некоторых нейронов.
Синапс
Переход возбуждения от нейрона к нейрону, межнейронное взаимодействие происходит посредством
синапсов — соединений, осуществляющих передачу специфических сигналов. Представление о синапсах связано с Шеррингтоном (1935), высказавшим предположение о существовании специальных структурно-функциональных образований, обеспечивающих контакты между нейронами.
Особенности рефлекторных реакций и некоторые свойства нервных центров обусловлены процессами, происходящими на синапсах. Синапс включает в себя три компонента: пресинаптический, постсинаптичес-кий и синаптический, т.е. содержит элементы и первого и второго контактирующих нейронов. Пресинап-тическая и постсинаптическая части разделены си-наптической щелью. Контактировать между собой могут разные части нейронов: чаще встречаются синапсы аксодендритные и аксосоматические, реже -сома-соматические, дендро-дендритные и дендро-со-матические. Существуют синапсы с химическим и электрическим способами взаимодействия между контактирующими нейронами.
Химические синапсы -- это преобладающий тип синапсов в мозгу млекопитающих и человека. В них пресинаптическая часть представлена утолщением терминали аксона в виде бутонов, внутри которых содержится множество круглых или овальных везику-лов диаметром от 20-40 до 120 нм. Внутри везикулов содержится химическое вещество — медиатор, участвующий в синаптической передаче. Медиатор выделяется пресинаптическим окончанием, проходит через синаптическую щель и, действуя на постсинапти-ческую мембрану, изменяет ее проводимость (рис. 2.3). Выделение медиатора в синаптическую щель происходит вследствие деполяризации пресинаптической мембраны приходящими по аксону потенциалами действия. При деполяризации пресинаптической мемб-

|
Рис. 2.3. Строение химического синапса: а — схема простого контакта; б — схема пластинчатого синапса (Шеперд, 1987)
раны открываются каналы для Са++, который, входя внутрь, способствует слиянию везикулов с мембраной; затем происходит процесс, аналогичный экзоцитозу. Количество выделяемого медиатора контролируется величиной деполяризации. Молекулы медиатора выделяются квантами: один квант — это содержимое одной везикулы. В качестве медиаторов синаптической передачи сегодня известно большое число химических веществ, которые разделяют на 4 основные группы:
1. Ацетилхолин.
2. Катехоламины (дофамин, норадреналин, адре
налин, серотонин).
3. Аминокислоты (глицин, гамма-аминомасляная
кислота, глютамат, цистеин и др.).
4. Пептиды.
Первые две группы медиаторов синтезируются из циркулирующих в крови предшественников; аминокислоты и пептиды — результат длинных целей мозгового метаболизма, начинающегося от глюкозы. На всех пресинаптических окончаниях одного нейрона
выделяется медиатор единой химической природы. Между химической природой медиатора и знаком его синаптического действия нет однозначной зависимости: один и тот же медиатор может оказывать как возбуждающее, так и тормозящее действие. Знак синаптического действия определяется свойствами пост-синаптической мембраны, так как рецепторы постси-наптической мембраны могут разным образом реагировать с медиатором и контролировать проводимость разных ионных каналов.
Высвободившийся медиатор диффундирует через синаптическую щель, которая имеет ширину 20-30 нм и различную зону контакта. На постсинаптической мембране существуют активные зоны, содержащие молекулярные рецепторы. В результате взаимодействия медиатора с рецептором изменяется проницаемость определенных ионных каналов через мембрану, возникает ионный ток, который приводит к возникновению постсинаптического потенциала. При раскрытии каналов для Ма1, а также Са++ происходит деполяризация мембраны, возникает возбуждающий постсинаптический потенциал (ВПСП). При раскрытии каналов для СГ и К+ происходит гиперполяризация мембраны, возникает противоположный по знаку тормозной постсинаптический потенциал (ТПСП). Интенсивность ионных потоков через мембрану и величина постсинаптических потенциалов градуально изменяются в зависимости от силы пресинапти-ческого воздействия и количества выделившегося медиатора. После того, как медиатор подействовал на рецепторы постсинаптической мембраны, синаптичес-кая щель очищается от медиатора путем его дезактивации или гидролиза, захвата глиальными клетками или пресинаптическим нейроном. Химические синапсы отличаются полярностью организации, односторон-
ним проведением, наличием синоптической задержки и химической чувствительностью постсинапти-ческой мембраны. Если активная мембрана аксона и сомы изменяет ионную проницаемость под действием электрического тока и способна генерировать распространяющиеся потенциалы действия, подчиняющиеся закону «все или ничего», то постсинаптическая мембрана изменяет ионную проницаемость в результате химического взаимодействия медиатора и рецепторов, генерирует неспособные к распространению постсинаптические потенциалы, амплитуда которых градуально изменяется, а они сами способны суммироваться.
Электрические синапсы широко распространены в нервной системе беспозвоночных и низших позвоночных животных. В стволе мозга млекопитающих они имеются в ядрах тройничного нерва, в вестибулярных ядрах Дейтерса и в нижней оливе мозгового ствола. В электрических синапсах узкие щелевые контакты отличаются низким электрическим сопротивлением, в них почти нет токов утечки через внеклеточную среду, поэтому изменения потенциала в пресинаптической мембране могут эффективно передаваться на электрочувствительную постсинапти-ческую мембрану, которая под воздействием потенциалов действия пресинаптической мембраны изменяет ионную проницаемость и может генерировать потенциалы действия. В ряде электрических синапсов потенциал действия передается с пресинаптической мембраны на постсинаптическую с меньшими потерями, чем в обратном направлении. В электрических синапсах проведение возбуждения происходит почти без еинаптической задержки, ток возможен в обоих направлениях, но легче в одном (эффект выпрямления); они дают возможность получать
постоянные, повторяющиеся реакции и синхронизировать активность многих нейронов.
2,4. Нейроглия
Наряду с нейронами нервная система содержит клетки нейроглии. В головном мозгу их существенно больше, чем нервных клеток: в некоторых отделах почти в 10 раз. Эти количественные соотношения свидетельствуют о важном значении глиальных клеток в физиологии нервной системы. Термин «глия» означает «связующее» и отражает роль нейроглии как посредника между кровеносными сосудами и нейронами. Клетки нейроглии неоднородны по своему происхождению: большинство видов глии возникает из нервной части эктодермы. Различают периферическую нейроглию, к которой относятся швановские клетки, и центральную нейроглию, состоящую из астро-цитов, олигодендроглии и микроглии.
Швановские клетки образуют оболочки периферических аксонов. Спиральное закручивание мембраны швановской клетки в процессе эмбрионального раз вития образует вокруг аксонов у позвоночных плотную складчатую изоляцию — миелиновую оболочку. Одна швановская клетка покрывает миелином 1 мм аксона. Миелиновая оболочка обеспечивает быстрое эффективное распространение возбуждения на большие расстояния, изоляцию аксонов друг от друга и выполняет вспомогательную функцию в отношении обмена веществ прилегающих нейронов.
Астроциты составляют около 25% всех центральных глионов и распределены в головном мозгу равномерно. Для астроцитов характерно большое количе ство лучеобразно расходящихся отростков, придающих им вид звезды; часть отростков заканчивается на поверхности кровеносных сосудов. За счет отроет-
ков астроциты имеют большую поверхность контактов и, соединяясь между собой, образуют пространственную сеть. У астроцитов существует несколько видов соединений, связывающих их друг с другом и с нервными клетками. Астроциты выполняют следующие функции: 1) служат каркасом для нервных клеток; 2) участвуют в метаболических процессах, влияющих на ионный состав и медиаторы синапти-ческой передачи; 3) обеспечивают репарацию нервов после повреждения; 4) изолируют и объединяют нервные волокна и окончания.
Олигодендроглия составляет 50-70% центральной нейроглии. Морфологически клетки отличаются малым количеством отростков, которые слабо ветвятся и значительно короче, чем у астроцитов. Цитоплазма олигодендроцитов характеризуется высокой метаболической активностью, содержит большое количество митохондрий и окислительных ферментов. Клетки олигодендроглии на больших поверхностях непосредственно контактируют с мембранами тел нервных клеток. Их функции связаны с осуществлением сложного метаболического обмена с нейронами и образованием миелина на мембране аксонов центральных нейронов.
Клетки микроглии имеют мезодермальное происхождение, это самые мелкие глиальные клетки, равномерно рассеянные по ЦНС. В сером веществе они расположены между отростками нейронов, нередко находятся в позиции сателлитов нервных клеток. Для микроглии характерна большая подвижность и способность фагоцитировать продукты распада нервных клеток. При воспалениях и повреждениях они движутся к очагу и, превращаясь в крупные макрофаги, выполняют роль «уборщика», слу-
жат защитой против воспаления и инфекции. Клетки микроглии выполняют сторожевую функцию, обходя нервную ткань, чтобы ликвидировать любые возникающие повреждения.
Электрические особенности нейроглии заключаются в том, что величина мембранного потенциала у них больше (90 мв), чем у нейрона (70 мв), и поэтому повышение концентрации К1 во внеклеточной среде приводит к деполяризации глиальной мембраны раньше, чем нервной. Глиальная мембрана является активным калиевым электродом. Мембраны глиальных клеток пассивно реагируют на электрический ток: их мембрана, в отличие от мембраны нейронов, не способна генерировать распространяющиеся потенциалы действия. Электрическое сопротивление глиальной мембраны (300 Ом/см2) ниже, чем нервной (5 000 Ом/см2). Между клетками глии существуют участки низкоом-ной связи, которые обеспечивают обмен ионов и мелких молекул.
Подводя итог, можно выделить следующие функции нейроглии: опорную, изоляционную, дренажную, транспортную, регулирующую ионный состав межклеточного пространства и объединяющую нейроны в группы. О функциональном значении глии свидетельствуют факты увеличения количества глиальных клеток вокруг активно функционирующих нейронов. Кратковременное возбуждение нейронов приводит к увеличению вокруг них глии за счет ее перемещения; длительные возбуждения нейронов вызывают увеличение глии за счет процессов клеточного деления. Предполагают, что глия, влияя на эффективность синаптической передачи, может участвовать в механизмах формирования временных связей при образовании условных рефлексов.
5. иощие свойства нервной системы
3.1. Эволюция нервной системы
В развитии нервной системы (НС) многоклеточных принято выделять три этапа, или три типа нервной системы - - диффузную (кишечнополостные), узловую (членистоногие) и трубчатую (позвоночные).
Эволюция нервной системы, ее структура и функции, как считает Сепп, должны рассматриваться в неразрывной связи с эволюцией моторики. И действительно, именно с этих позиций можно объяснить и структурные и функциональные преобразования нервной системы от кишечнополостных до высших позвоночных животных. Движение гидры напоминает амебоидные — в моторике участвует все тело, характер его перистальтический. В каком бы участке тела ни возникло возбуждение (например, при механическом раздражении), в этот процесс вовлекается вся нервная система, и гидра дает на это стереотипный ответ - - тотальное сокращение всей мускулатуры. Вторая ступень моторики — выделение специализированных частей тела, обеспечивающих передвижение (жгутики, реснички). Характер движения сохраняется прежний — перистальтический, бесскелетный, координация двигательной функции осуществляется нервным аппаратом, заключенным в самом двигательном органе. В наиболее чистом виде эта ступень представлена у мягкотелых. Коренное преобразование моторики связано с развитием скелета (третья ступень). В этом случае речь идет о движении с помощью рычагов. Рычаговое движение развилось у животных в двух вариантах, характеризующих членистоногих и позвоночных животных. У первых моторный скелет является одновременно и защитным (эк-зоскелет), у вторых он оказался внутри мышц, при-
водящих его в движение (эндоскелет). Рычаговая форма моторики потребовала чрезвычайного усложнения управляющего аппарата -- нервной системы, ввиду бесконечного числа степеней свободы изменения соотношений частей тела.
Эволюцию структуры и функции нервной системы следует рассматривать как с позиции совершенствования отдельных ее элементов — нервных клеток, так и с позиции совершенствования общих свойств, обеспечивающих приспособительное поведение.
Первым этапом развития НС было формирование диффузной НС. Нервные клетки диффузной НС мало напоминают нейроны позвоночных. В частности, в них отсутствует субстанция Ниссля, ядро не дифференцировано, количество отростков невелико, их длина незначительна. Передача возбуждения от нейрона к нейрону осуществляется не только синаптическим путем, но и через посредство протон лазматических мостиков. Нейроны слабо дифференцированы по функции. В частности, у гидроидов описаны так называемые нервно-сократительные элементы, где соединена функция нервных и мышечных клеток. В диффузной нервной системе имеются не только «локальные нервные» сети, образованные короткоотростчатыми нейронами, но и «сквозные пути», проводящие возбуждение на сравнительно большое расстояние. Скорость распространения возбуждения по волокнам много ниже, чем у других животных, и составляет сотые и десятые доли метра в секунду (табл. 3.1). При этом следует отметить, что низкая скорость передачи управляющих команд обусловлена также и необходимостью многократного переключения в х*епях корот-коотростчатых элементов. Диффузная нервная система, как уже указывалось, характерна для типа кишечнополостных. Этот тип включает в себя разные
формы — прикрепленные к субстрату (неподвижные) и ведущие свободный образ жизни. И хотя и в первом, и во втором случаях тип нервной системы характеризуется как диффузный, свойства и нейронов и нервных сетей могут различаться весьма значительно. Это еще раз подтверждает роль моторики в организации нервной системы. У свободноплавающих медуз появляются скопления нервных клеток (прототип нервных центров) в колоколе — краевые органы, выделяются сквозные проводящие пути, обеспечивающие определенную «адресность» в проведении возбуждения. Основной особенностью диффузной нервной системы является неопределенность связей, отсутствие четко выраженных входов и выходов, надежность функционирования, но энергетически эта система малоэффективна.
Таблица 3.1
Date: 2016-11-17; view: 385; Нарушение авторских прав