Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Подтип Трахейнодышащие (Tracheata) 6 page
|
|
Взаимодействия аллельных генов. Явление, когда за один признак отвечает несколько генов (аллелей), называется взаимодействием генов. Если это аллели одного и того же гена, то такие взаимодействия называются аллельными, а в случае аллелей разных генов —неаллельными.
Типы аллельных взаимодействий: доминирование, неполное доминирование, сверхдоминирование и кодоминирование.
Доминирование - тип взаимодействия двух аллелей одного гена, когда один из них полностью исключает проявление действия другого: 1) доминантный аллель в гетерозиготном состоянии обеспечивает синтез продуктов, достаточный для проявления признака такого же качества, как и в состоянии доминантной гомозиготы у родительской формы; 2) рецессивный аллель совсем неактивен, либо продукты его активности не взаимодействуют с продуктами активности доминантного аллеля. (пурпурная окраска цветков гороха над белой, гладкая формы семян над морщинистой, темные волосы над светлыми, карие глаза над голубыми у человека и т. д.
Неполное доминирование, или промежуточный характер наследования, наблюдается в том случае, когда фенотип гибрида (гетерозиготы) отличается от фенотипа обеих родительских гомозигот, т. е. выражение признака оказывается промежуточным, с большим или меньшим уклонением в сторону одного или другого родителя. Механизм этого явления состоит в том, что рецессивный аллель неактивен, а степень активности доминантного аллеля недостаточна для того, чтобы обеспечить нужный уровень проявления доминантного признака. Пример: наследование окраски цветков у растений ночной красавицы. При скрещивании растения с красными цветками и растения с белыми цветками в F1, у всех растений цветки розовые, т. е. наблюдается промежуточный характер наследования. При скрещивании гибридов с розовой окраской цветков в F2 имеет место совпадение расщепления по фенотипу и генотипу, так как доминантная гомозигота (АА) отличается от гетерозиготы (Аа). Так, в рассматриваемом примере с растениями ночной красавицы расщепление в F2 по окраске цветков обычно следующее — 1 красная (АА): 2 розовые (Аа): 1 белая (аа).
Также: курчавость волос у человека, масть КРС, окраска оперения у кур.
Сверхдоминирование — более сильное проявление признака у гетерозиготной особи (Аа), чем у любой из гомозигот (АА и аа). Предполагается, что это явление лежит в основе гетерозиса.
Кодаминирвание— участие обоих аллелей в определении признака у гетерозиготной особи: наследование IV группы крови у человека (группа АВ).
Взаимодействия неаллельных генов.Неаллельные взаимодействия генов описаны у многих растений и животных. Они приводят к появлению в потомстве дигетерозиготы необычного расщепления по фенотипу: 9:3:4; 9:6:1; 13:3; 12:3:1; 15:1, т.е. модификации общей менделевской формулы 9:3:3:1. Известны случаи взаимодействия двух, трех и большего числа неаллельных генов. Среди них можно выделить следующие основные типы: комплементарность, эпистаз и полимерию.
Комплементарным, или дополнительным, называется такое взаимодействие неаллельных доминантных генов, в результате которого появляется признак, отсутствующий у обоих родителей. Например, при скрещивании двух сортов душистого горошка с белыми цветками появляется потомство с пурпурными цветками. Если обозначить генотип одного сорта ААbb, а другого — ааВВ, то
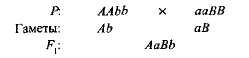
Гибрид первого поколения с двумя доминантными генами (А и В) получил биохимическую основу для выработки пурпурного пигмента антоциана, вто время как поодиночке ни ген А, ни ген B не обеспечивали синтез этого пигмента. Синтез антоциана представляет собой сложную цепь последовательных биохимических реакций, контролируемых несколькими неаллельными генами, и только при наличии как минимум двух доминантных генов (А-В-) развивается пурпурная окраска. В остальных случаях {ааВ- и A-bb) цветки у растения белые.
При самоопылении растений душистого горошка из F1 в F2 наблюдалось расщепление на пурпурно- и белоцветковые формы в соотношении, близком к 9:7. Пурпурные цветки были обнаружены у 9/16 растений, белые — у 7/16.
Эпистаз — это такой тип взаимодействия генов, при котором аллели одного гена подавляют проявление аллельной пары другого гена. Гены, подавляющие действие других генов, называются эпистатическими, ингибиторами или супрессорами. Подавляемый ген носит название гипостатический. Доминантный эпистаз (А>В или В>А) с расщеплением 12:3:1; рецессивный эпистаз (а>В или b>А), который выражается в расщеплении 9:3:4, и т. д.
Полимерия проявляется в том, что один признак формируется под влиянием нескольких генов с одинаковым фенотипическим выражением. Такие гены называются полимерными. В этом случае принят принцип однозначного действия генов на развитие признака. Например, при скрещивании растений пастушьей сумки с треугольными и овальными плодами (стручочками) в F1 образуются растения с плодами треугольной формы. При их самоопылении в F2 наблюдается расщепление на растения с треугольными и овальными стручочками в соотношении 15:1. Это объясняется тем, что существуют два гена, действующих однозначно. В этих случаях их обозначают одинаково— А1и A2.
Тогда все генотипы (А1,-А2,-, А1-а2а2, a1a1A2-) будут иметь одинаковый фенотип — треугольные стручочки, и только растения а1а1а2a2 будут отличаться —- образовывать овальные стручочки. Это случай некумулятивной полимерии.
Полимерные гены могут действовать и по типу кумулятивной полимерии. Чем больше подобных генов в генотипе организма, тем сильнее проявление данного признака, т. е. с увеличением дозы гена (А1 А2 А3 и т. д.) его действие суммируется, или кумулируется. Например, интенсивность окраски эндосперма зерен пшеницы пропорциональна числу доминантных аллелей разных генов в тригибридном скрещивании. Наиболее окрашенными были зерна А1А1А2А2А3,А 3 а зерна а1а1а2a2а3а 3 не имели пигмента.
Также: молочность, яйценоскость, масса и другие признаки сельскохозяйственных животных; многие важные параметры физической силы, здоровья и умственных способностей человека; длина колоса у злаков; содержание сахара в корнеплодах сахарной свеклы или липидов в семенах подсолнечника и т. д.
Проявление большей части признаков представляет собой результат влияния комплекса взаимодействующих генов и условий внешней среды на формирование каждого конкретного признака.
4.Типы взаимодействия аллельных и неаллельных генов, их молекулярные механизмы.
Различают следующие основные типы взаимодействия генов:
1) комплементарность; 2) элистаз; 3) полимерия; 4) модифицирующее действие генов.
Комплементарное действие генов. Гены называют комплементарными, когда они по своему проявлению как бы дополняют друг друга. Каждый такой ген в отдельности, сам по себе, не оказывает действия, не вызывает появления признака. Однако при скрещивании двух особей, из которых каждая имеет такого типа ген (например, одна имеет ген А, а другая ген В), получается гибрид, новый организм, у которого в наличии оказываются оба этих гена, и эффект их совместного действия проявляется в том, что под их совместным влиянием у гибридного организма возникает какой-то новый признак. Например, Куры с розовидным гребнем и меют генотипАвв, с гороховидным – ааВ, с листовидным гребнем – аавв. При скрещивании кур с розовидным и гороховидным гребнем получается новый признак – ореховидный гребень.
Эпистатическое действие генов (эпистаз)
Эпистатическое действие генов по своему характеру противоположно комплементарному действию генов. Сущность его состоит в подавлении генами супрессорами действия гипостатических генов. Явление эпистаза выражается в неаллельном подавлении действия одного доминантного гена другим доминантным геном, принадлежащим к другой аллеломорфной паре.
Гены, которые подавляют действие других неаллельных им (принадлежащих к другим парам) генов, называют эпистатичными.
Эпистаз выражается в изменении соотношения расщепления во втором поколении, которое по фенотипу отклоняется от обычного расщепления при дигибридном скрещивании, т. е. от нормы Менделя 9:3:3: 1.
Рассмотрим явление эпистаза схематически. Существуют два разных по окраске типа лошадей: доминантный ген С характеризует серую окраску, а доминантный ген В — черную, вороную.
При скрещивании генотипы родителей будут: ССвв (серая) × ссВВ (вороная), а генотип F1 CcBe.
Окраска потомства F1 будет серая, так как ген С (доминант) эпистатически подавляет проявление гена В вороной окраски.
Скрестив между собой эти генотипы, т. е. Сс × Вв, будем иметь в F2 расщепление по фенотипам, 12: 3: 1.
Мы видим, что: 1) все зиготы, имеющие доминантный ген С, дают серых лошадей, так как ген С серой окраски эпистатичен доминантному гену В вороной окраски; 2) все зиготы с рецессивом с и доминантом В дадут вороную окраску, так как рецессивный ген с не подавляет действия доминантного гена В; 3) двойной рецессив ссвв т. е. форма, гомозиготная по обоим рецессивным генам, дает форму, отличную по фенотипу от форм с доминантными аллелями двух генов С и В, а также от форм с одной из таких аллелей (ссВВ, ССвв). В данном случае это отличие по фенотипу выражено в рыжей окраске лошади.
Полимерное действие генов (полимерия)
Полимерия представляет собой явление взаимодействия генов, при котором несколько однотипных (однозначных) генов оказывают сходное воздействие на развитие одного и того же признака.
Иначе говоря, полимерия обусловливается действием разного числа однозначных генов, которые, суммируясь, усиливают проявление признака, а при меньшем числе таких генов этот признак проявляется в соответственно меньшей степени.
С явлением полимерии приходится сталкиваться при изучении так называемых количественных признаков.
Такие признаки, как, например, вес животного, яйценоскость кур, количество белка, в эндосперме зерна кукурузы и зерна пшеницы, содержание витаминов в растениях, скорость протекания биохимических реакций и т. п., нельзя разложить на четкие фенотипические классы; их необходимо оценивать и измерять количественно. Такие признаки называются количественными, или мерными.
Изучение наследования количественно варьирующих признаков у различных особей одного и того же поколения было начато в первом десятилетии XX в. Шведский генетик Нильсон-Эле в 1908 г., скрещивая расы пшеницы, имеющие красные и белые зерна, обнаружил в первом поколении F1 обычное моногибридное расщепление 3:1. Однако при скрещивании некоторых линий пшениц с такими признаками он во втором поколении (F2) в 1910 г. получил расщепление в соотношении 15/16 окрашенных и 1/16 белых. Окраска зерен у первой группы, т. е. у 15/16 растений, варьировала от темно-красной до бледно-красной.
По типу полимерии, т. е. полимерных генов, наследуется цвет кожи у человека. Так, от брака негра и белой женщины рождаются дети с промежуточным цветом кожи (мулаты). А у супружеской пары двух мулатов рождаются дети всех возможных типов при комбинации двух неаллельных полимерных генов - от черной до белой кожи.
Изучение полимерных (множественных) генов имеет большое значение, так как очень многие хозяйственно ценные признаки у растений и животных наследуются по типу полимерии (например, содержание сахара в корнеплодах свеклы, длина початка кукурузы и т. д.).
Date: 2015-09-24; view: 578; Нарушение авторских прав