Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Подтип Трахейнодышащие (Tracheata) 1 page
|
|
Высшие трахейнодышащие членистоногие. Дышат только с помощью трахей. Имеется одна пара усиков, которая гомологична антеннулам (1-й паре антенн) ракообразных и принадлежит акрону. Сердце трубчатое, располагается на спинной стороне полости тела. Органы выделения -мальпигиевы трубочки. Подразделяется на 2 класса.
Класс 1. Многоножки (Myriapoda)
Низшие трахейные. Тело подразделено на 2 отдела - голову и туловище, состоящее из многих сегментов. Крыльев нет. Эпикутикулы (самый наружный, очень тонкий, влагонепроницаемый слой кутикулы) нет. Ведут скрытный образ жизни. Днем прячутся под корой, стволами деревьев и в других укрытиях.
Класс 2. Насекомые (Insecta)
Высшие по своей организации и степени освоения различных сред обитания членистоногие и вообще первичноротые. Крупный ароморфоз -приобретение в процессе эволюции насекомыми крыльев позволил им освоить воздушное пространство как новую среду обитания, без утраты наземной сферы как среды обитания. Тело состоит из трех отделов -головы, груди, брюшка. Строение ротового аппарата очень разнообразно и связано с характером пищи и способом ее приема. На груди располагаются 3 пары ног, имеющих различные модификации в связи с характером передвижения и среды обитания. Крыльев две или одна пара крыльев, которые нередко частично или полностью редуцированы в связи со специфическим образом жизни, например, у эктопаразитов. Самый наружный слой кутикулы - эпикутикула (сложный комплекс восков и липидов), хорошо защищающий организм от потери воды. Особенности ротового аппарата и всей пищеварительной системы связаны с характером питания. Органы дыхания - сложно разветвленная трахейная система. Развитие у большинства с неполным или полным превращением. Играют огромную роль в природе как опылители растений, потребители живой и мертвой органической продукции, мелиораторы почвы. Для человека полезны опылением культурных и дикорастущих покрытосеменных растений, как поставщики ценных пищевых продуктов и лекарственных материалов (мед, прополис и их производных, натуральный шелк), как регуляторы численности вредных видов (паразиты и хищники). Ряд видов насекомых - вредители сельского и лесного хозяйства (клоп вредная черепашка, колорадский жук, хлебный жук кузька, бабочки капустная белянка, луговой мотылек, озимая совка и др.).
20.Насекомые как высшие трахейнодышащие членисnоногие. Ароморфозы, позволившие им широко освоить наземную и воздушную среды. Важнейшие черты организации. Классификация, разнообразие, экологические адаптации, роль в природе и для человека.
Насекомые (Insecta) двустороннесимметричные, трехслойные, первичноротые, с сегментированным телом, гетерономной метамерией. Покровы - хитиновая кутикула; конечности членистые функции: локомоторную, защитную, органов схватывания и измельчения пищевых материалов, половых придатков. Полость тела смешенная – миксоцель; кровеносная система незамкнутая; рост сопровождается линьками. При этом ароморфозом – гетерономную сегментацию, членистые конечности и их прогрессивную дифференцировку в процессе адаптивной радиации, включая дифференцировку ротового аппарата. Приобретение в процессе эволюции крыльев) позволило крылатым насекомым освоить воздушное пространство как новую среду обитания. Тело состоит из 3 отделов - головы, груди, брюшка. Скрыточелюстные (Entognata)-грудной отдел слабо обособлен от брюшного. Есть мандибулы и максилы. На голове усики, простые глазки, на брюшке - двигательные, дыхание трахейное или только кожное. отр Бестяжковые, отр. Ногохвостки, отр. Двухвостки. Насекомые открыточелюстные (Entognatha) – тело состоит головы с усиками и 3 пар ротовых конечностей, брюшка без конечностей; дыхание трахейное. Достигают прогресса благодаря жизни на суши (строение покровов, многофункц особенности выделительной системы, трахейной, появление органов полета, особ эмбриогенеза и постэмбриогенеза обеспечивают независимость от факторов сухопутной среды. Пищеварительная система состоит из 3 отделов, есть слюнные железы, пищеварительные ферменты. Органы выделения: мальпигиевые сосуды (осморегуляции, накопления) Кров.система - незамкнутая, кровь – гемолимфа (снабжение органов питат вещ-вами,гуморальная регуляция), есть сердце. Дыхат: трахеи и дыхальца.(транспорт О2 к тканям). Н.С.: брюшная нервная цепочка, есть г.м. С ЦНС связана сист. симпатических нервов. Размножение половое, раздельнополые; диморфизм, партеногенез. развитие с превращением или нет. П.кл. Первичнобескрылые - низкие черты организации крыльев. Рот.аппарат - грызущий; раз - прямое. Отр.Щетинохвостки. П.кл. Крылатые (Pterygota): Поденки (Ephemeroptera), стрекозы (Odonata), Таракановые (Blattodea), Богомоловые (Mantodea), Термиты (Isoptera), Прямокрылые (Orthoptera), Вши (Anoplura), Равнокрылые (Homoptera), Клопы (Hemiptera), Сетчатокрылые (Neuroptera), Жуки (Coleoptera), Перепончатокрылые (Hymenoptera), Двукрылые (Diptera), Блохи (Aphaniptera), Ручейники (Trichoptera), Бабочки (Lepidoptera). Значение: важное звено пищевых цепей; производят полезные для человека продукты (мед, воск, прополис, пчелиный яд (медицина); объекты биотехнологии (для борьбы с нас вредителями).
21.Общая характеристика иглокожих как вторичноротых древних морских животных, обладающих вторичной радиальной симметрией. Особенности организации и образа жизни морских звёзд, морских ежей и голотурий.
Тип Иглокожие(Echinodermata) составляют морские целомические животные; вторичноротые. Распространены во всех соленых морях на разных глубинах. Сочетание билатеральной симметрии с радиальной(5-лучевой). Обладают известковым скелетом (функция: защита, опора для внутренних органов). Вторичная полость тела - целом, выполняет функции: полость для органов, гомеостаз внутренней среды, опора, транспорт. Характерна амбулакральная система – двигательная, на амбулакральных ножках, которые служат еще и для подачи пищи ко рту. Н.с. неганглиозного типа, закладывается в виде тяжа путем погружения внутрь зародыша эктодермальной пластинки. Дыхание через кожу, кожными жабрами, амбулакральными ножками, щупальцами. Органов выделения нет. Амебоцисты поглощают из целома экскреты, которые затем удаляют через кожу. Н.с. состоит из нервного кольца и отходящих от нее радиальных нервных стволов. Органы чувств: чувствительные клетки; равновесия (глубоководных голотурий). Раздельнополый деморфизм выражен слабо. Оплодотворение наружное; личинка – диплевра. Есть живородящие. 4 типа питания: зоо-, фито-, детрито-,планктонофагия. Кл. Морские звезды (Asteroidea)- звездообразные формы. Различные типы питания, движения. Особенности – наличие радиальных лучей, в которые заходят органы. Характерно скользящие движение амбулакральных ножек. (Asterias rubens) Кл. Морские ежи (Echinoidea) Малоподвижные, донные, шаровидной или яйцевидной формы. Хорошо развит скелет, образующий панцирь; движутся на амбулакральных ножках. 2 Подкласса: Правильные ежи и Неправильные ежи. У правильных наличие жевательного аппарата - Аристотелев фонарь. Личинка - эхиноплетеус. Кл. Голотурии (Holothuroidea) Медленно ползают; тело продолговатое; билатеральная симметрия преобладает; детритофаги; раздельнополые, есть гермафродиты.
Роль: в биологических цепях, в почвообразовании, в состав осадочных пород - мрамор, известняк; голотурии в пищу.
22.Позвоночные. Череп и его эволюционные преобразования в ряду позвоночных. Черты прогресса и адаптивные особенности черепа позвоночных.
Развитие головного мозга и связанных с ним органов чувств требовало образования для их защиты новых скелетных образований. Эту функцию выполняет мозговой, или осевой череп. К тому же, переход к активному питанию подвижными пищевыми объектами вызывал необходимость образования дополнительных органов активного захвата и переработки пищи. Это обеспечено формированием висцерального скелета, окружающего передний конец пищеварительной трубки, а у водных позвоночных этот отдел служит и опорой жабр. Висцеральный (splanchnocranium) и мозговой (neurocranium) череп у всех позвоночных животных объединяется в единый комплекс - череп (cranium). Впервые в эволюционном ряду позвоночных он начинает формироваться у круглоротых.
Основные этапы эволюционные преобразования черепа наглядно прослеживаются при рассмотрении эмбрионального развития позвоночных.
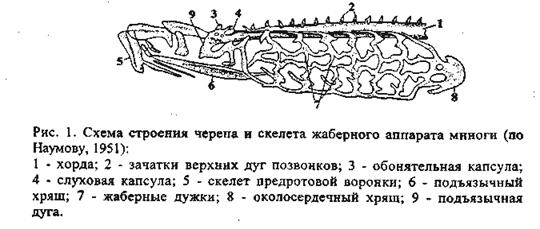
Мозговой череп закладывается под головным мозгом в виде двух-трех пар хрящей. Одновременно, но независимо от них возникают хрящевые капсулы органов чувств: обонятельные, слуховые и глазные. Разрастаясь и сливаясь друг с другом, они начинают обрастать головной мозг с боков. Такая стадия развития мозгового черепа, еще лишенного крыши, соответствующая ранним этапам эмбрионального развития черепа остальных позвоночных, присуща круглоротым (рис. 1).
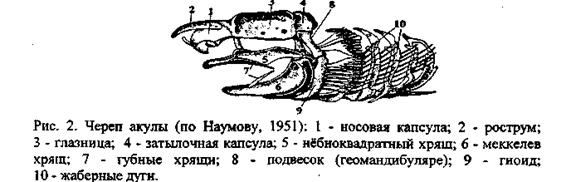
У остальных позвоночных еще на личиночной или зародышевой стадии разросшийся хрящ образует и крышу черепа. Такой, полностью хрящевой череп, у хрящевых рыб - акул и скатов (рис. 2).
У низших костных рыб (хрящевых ганоидов) мозговой череп остается хрящевым, а поверх него образуется панцирь из покровных кожных костей (рис. 3). У остальных групп костных рыб в первичном хрящевом мозговом черепе возникают окостенения, завершающиеся образованием так называемых первичных, или хрящевых, костей. Одновременно покровные кости погружаются под кожу и вступают в соединение с хрящевыми костями. У костистых рыб мозговой череп состоит из большого количества хрящевых и покровных костей; хрящ сохраняется только в области обонятельных и слуховых капсул и глазниц (рис. 4).
У современных амфибий в мозговом черепе остались значительные участки хряща и относительно небольшое число хрящевых и покровных костей (рис. 5).

|

|

У рептилий, птиц и млекопитающих мозговой череп во взрослом:состоянии образован лишь костями. У птиц и млекопитающих большинство костей срастается между собой, так что их границ обнаружить нельзя. Впервые У некоторых пресмыкающихся, а позже и у остальных позвоночных, в мозговом черепе образуется вторичное твердое костное нёбо, разделяющее полость рта на верхний отдел - носоглоточный ход - и нижний отдел -собственно ротовую полость (рис. 6). Благодаря этому пища, находящаяся во рту, не мешает дыханию.
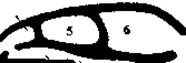
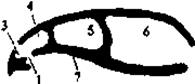
Рис. 6. Схема образования вторичного твердого костного нёба (по Шмальгаузену, 1947).
Первичные хоаны; 2 - вторичные хоаны; 3 - дыхательная и 4 - чувствующая части обонятельной полости; 5 - орбита; 6 - черепная полость; 7 - первичное нёбо; 8 - вторичное твёрдое нёбо; 9 - носоглоточный проход.
Эволюция жевательной мускулатуры, начиная с пресмыкающихся, сопровождалась перестройкой крыши и частично боковых стенок первично сплошного «стегального» черепа, имевшегося у древних земноводных -стегоцефалов, предков рептилий. В отдельных ветвях пресмыкающихся эволюция шла разными путями. В группе анапсида (ископаемые котилозавры и их потомки - черепахи) первичная стегальная (сплошная) крыша черепа сохранялась. В группе диапсида (гаттерия, крокодилы) образовались две височные ямы, ограниченные двумя костными дугами (рис. 7).
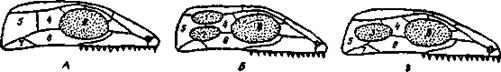
Рис. 7. Схема изменения строения черепа у пресмыкающихся (по Наумову и Карташеву, 1979):
А - стегальный, или анапсидный череп; Б - диапсидный череп с двумя височными ямами; В - синапсидный череп с одной боковой височной ямой. 1 - верхняя височная яма, 2 нижняя височная яма, 3 - единственная боковая височная яма, 4 — заглазничная кость, 5 - чешуйчатая кость, 6 - скуловая кость, 7 - квадратно-скуловая кость, 8 - орбита.
Верхняя дуга состоит из костей залобной (postfrontale) - чешуйчатой (squamosum), нижняя - из скуловой (]U8ale) - квадратноскуловой (quadratojugale). У ящериц диапсидный тип черепа с редуцированной нижней дугой, а у птиц - с редуцированной верхней дугой; у змей обе дуги редуцируются. В группе синапсид - у звероящеров (тероморфных рептилий) и происшедших от них млекопитающих - образовалась одна боковая яма и ограничивающая ее сложная дуга из костей верхней и нижней дуг: скуловой (jugale) - квадратноскуловой (quadratojugale) - чешуйчатой (squamosum).
По особенностям расположения мозга в черепной коробке различают два основных типа мозгового черепа. Платибазальный череп (круглоротые, хрящевые рыбы, амфибии) имеет широкое основание и мозговую полость, расположенную между его глазницами, а тропибазальный (костные рыбы, рептилии, птицы, млекопитающие) - узкое основание, глазницы, разделенные лишь тонкой межглазничной перегородкой, и мозговую полостью, размещенную позади глазниц (рис. 8).

По мере движения по эволюционному ряду позвоночных от костистых рыб к млекопитающим происходит постепенное уменьшение количества костей, образующих череп. Наименьшее количество их у млекопитающих.
Висцеральный отдел черепа развивается независимо от мозгового черепа в виде жаберных дуг, лежащих в перегородках между жаберными щелями. У круглоротых эти хрящевые зачатки преобразуются в хрящевую жаберную решетку околосердечный хрящ и в хрящи, поддерживающие язык и ротовую воронки.
У водных челюстноротых позвоночных животных рудименты первых двух жаберных дуг образуют губные хрящи (развиты у акуловых рыб). Третья дуга, которую называют челюстной, расчленяется на два отдела, образуя собственно челюсти: верхний элемент называют небно-квадратным хрящом (palatoquadratum), нижний - меккелевым хрящом (cartilage meckeli). Эти элементы выполняют функцию челюстей только у низших рыб (см. рис. 2).
У высших рыб они окостеневают и лишь частично участвуют в удержании добычи; а захват добычи у них переходит вторичными челюстями, образованными покровными костями: верхнечелюстной (maxillare) и предчелюстной (praemaxillare) - в верхней челюсти; зубной (dentale) - в нижней челюсти.
Следующая висцеральная дуга, подъязычная, обычно состоит из двух крупных парных хрящевых элементов: подвеска, или гиомандибулярного (hyomandibulare), и подъязычного, или гиоида (hyoideum). Гиоиды правой и левой сторон соединяются друг с другом при помощи небольшого непарного элемента - копулы (copulae). У высших рыб подъязычная дуга окостеневает (рис. 4).
Остальные дуги служат опорой жабр - жаберные дуги. У примитивных рыб число их может достигать 7 пар, у большинства хрящевых рыб - 5 пар хрящевых жаберных дуг, у костистых рыб часто их 4 пары (V-рудиментарна) и они окостеневают. За счет покровных костей у костных рыб формируется скелет жаберной крышки, тоже относящийся к висцеральному черепу.
Различают несколько типов прикрепления челюстного аппарата к мозговому черепу (рис. 9).

Рис. 9. Отношение передних висцеральных луг к осевому черепу (по Шмальгаузену, 1947).
А- гиостильный череп акулы; В - амфистильный череп акулы; С - аутостильный череп наземного позвоночного (лягушка); hm - hyomandibulare; hy - hyoideum; mk - меккелев хрящ; md - нижняя челюсть; ot - processus oticus; pb - processus palatobasalis; pq - palatoquadratum.
1. Протостилия - челюстная и подъязычная дуги независимо друг отдруга связками подвешивались к мозговому черепу. Этот гипотетический исходный тип черепа, вероятно, был свойствен примитивным челюстноротым.
2. Гиостилия - верхний конец гиомандибулярного отдела подъязычной дуги прикрепляется к слуховому отделу мозгового черепа и служит подвеском для челюстной дуги (многие группы рыб, в том числе большинство хрящевых и все костистые рыбы).
3. Амфистилия - верхний элемент челюстной дуги соединяется с мозговым черепом при помощи одного - двух специальных отростков и, кроме этого, как и при гиостилии, задние концы обоих челюстных элементов прочно связаны с нижним концом гиомандибулярного элемента (некоторые примитивные и современные акулы, костные ганоиды).
4. Аутостилия - верхний элемент челюстной дуги соединяется или срастается с мозговым черепом, гиомандибулярный элемент подъязычной дуги в прикреплении челюстей не участвует, и подъязычная дуга в большей или меньшей степени подвергается редукции (цельноголовые и двоякодышащие рыбы, земноводные и все другие наземные позвоночные).
С переходом к воздушному дыханию висцеральный скелет видоизменяется. В ряду земноводные - млекопитающие происходит усиление вторичных челюстей, а развитие аутостилии приводит к редукции подъязычной дуги: нижний ее элемент вместе с редуцирующимися жаберными датами участвует в образовании подъязычного аппарата и скелета гортани, а верхний элемент - гиомандибулярный, или подвесок - превращается в косточку среднего уха - стремечко. Скелет жаберной крышки полностью редуцируется.
У млекопитающих, в отличие от всех остальных позвоночных, нижнюю челюсть образует одна единственная парная кость - зубная (dentale).
23.Выделительная система в ряду хордовых. Особенности и направления её эволюции. Позвоночные. Способы поддержания гомеостаза. Терморегуляция. Осморегуляция.
Обеспечивает выделение жидкого конечного продукта обмена. Почки – главный орган выделения. Многие сталкиваются с проблемой осморегуляции, осморегуляция – один из параметров поддержания гомеостаза.
По величине осмотического давления растворы м.б. изотоническими (изоосмотич) – 2 р-ра им одинак давление, гипертонические – с более высок осмотич давл, гипотонич – с более низк. Водные организмы по этой величине м.б. изоосмотич, гипоосмотич.
В теле животных жидкости по своему состоянию напоминают морскую воду, они предст собой солев р-ры, электролит – NaCl, т.к. жидкость зародилась в море, организмы д были быть этой воде подобны. В то же время морская вода и жидкости тела не тождественны: и состав морск океана менялся, и состав жив менялся – проблема осморегуляции стоит и перед орг, жиб в воде. Морск орг: их жидкости менее концентрированы, чем окр среда – чистая вода поступает в тело. Удаление воды осущ ч/з жабры и почки. У морск рыбы чаще всего происх пассивн выд-е воды жабрами, нек кол-во изотонич мочи вывод ч/з почки. Пресноводные рыбы (и личинки амфибй): конц-ция вещ-в внутри тела намного превышает конц-цию вещ-в пресной воды. Пассивное поглощение воды телом и жабрами, почки выделяют оч большое кол-во гипотонич мочи с неб сод-ем солей.
Продукты экскреции: продукты распада АК и белков – соед-я, сод аминогр (токсичны) – выделение аммиака (м. олько круглоротые, рыбы и личинки аф) – амониотелия. Аммиак мгновенно растворяется в окр воде, легко вымывается, жабры выд аммиак, избыток аммиака накапл в мышцах. У др животных аммиак превращ-ся в мочевину (менее токсичное соед-е, но ещё ядовита и требует разведения). Выд-е мочевины – уреотелия. Мочевина растворима в воде – продукты выд-я оказ жидкими – выд достаточно больш кол-во мочи, осмотич св-ва м.б различны. Уреотелия: взрослые амф, млекопит. Мочевина обр в печени, поступает в кровь, ыд ч/з почки. У нек выд-ся мочевая к-та – урикотелия. Моч к-та не раств в воде, продукты выд сухие, мочи выд мало, но она высок конц – рептилии, птицы. Уринотелия – выд-е уринов (насек, паукообр) не св-на позвоночным.
У хордовых функ 2 осн гр орг:
Низшие – метанефридии – воронки с булавовидн обр со жгут – соленоциты.
Черепные – почки. Строение почек различно. Хар-на смена 3х поколений почек: в эмбриональном развитии набл 3 пок: предпочка (головная почка – пронефрос) – имеются внешние мальпигиевы клубочки, расп в целомич жидк, мальп кл – кап сеть, СОБР в клубок, со ст выд сист имеются воронки, откр в целом, имеется первичный мочеточник – пронефрос (пронефрич канал = вольфов канал – стекание мочи, выводн проток выделит сист). Сам перв почка обособл – воронка яйцевода, от неё отх ещё один канал – мюллеров. Им воронки с ресничками – нефростомы, кр сист обл мальп клубочками, прям контакта м/у кров и выд сист нет (обмен ч/з целомич полость). Пронефрос заклад-ся у эмбрионов практич у всех позвоночных, но функц только у нек круглоротых.
2е пок-е почек заклад-ся ниже – туловищная почка =первичная почка, мезонефрос). Имеется непосредств контакт кр и выд сист, жидк филтр непоср ч/з клубочки. Внтур мальп клубочки = мальп тельца. Мальп кл окружается капсулой (капсула Боумена) мальп клуб + капс Боумена место фильтрации) = мальп тельце.
У сам примит сох воронки (хрящ рыбы), в кач-ве мочеточников функц-ет Вольфов канал. У самок хорошо выражен Мюллеров канал, кот служит для выд-я прод. Тул почка функц-ет у амфибий, у хрящ и костн рыб.
У амниот (рептилии, птицы, млекопит) в эмбриональн развитии функц-ет первичн поки – мезонефросы, во взр сост ф-ет 3е пок-е почек – вторичные почки – метанефрос (тазовые почки). Развитие нефронов. Нефрон – структурно-функц ед почки. Нефрон предст собой мальп тельце, здесь происх фильтрация под давлением +в нефр им извитые канальцы (сост из канн 1го и 2го порядка). В обл канальцев им каппилярн сеть (активн экскреция – обр из первичн мочи вторичн моча, обратное всасывание воды в кровь и нек вещ-в, не подл выд-ю: глюкоза, нек соли), от метанефроса отщепл отдельн выделит канал – вторичн мочеточник, служит для выд-я конечн продукта обмена.
Рыбы, амфибии: Вольф канал – выд-е мочи и пермы. У самок от вольф канн – мюллеров канн, функц только в кач яйцевода. Выд-е ч/з вольфов канал.
Рептилии, птицы: у самцов обр вторичн почка, вторичн мочеточник, кот вп в остатки вольфова канала, вольф канал превращ в семепровод, в нижн отд происх соед-е – прод выд-я и пол прод ч/з 1 канал. Самки: вторичн мочеточник откр сам-но. Половой канал – мюллеров канал (яйцевод)
Млекопит: не им клоаки, хорошо развиты первичн пол органы.
Эволюция мочепол сист сводится к:
-смена 3х поколений поек: головн – туловищн – таз
-происх разделение каналов, лужащих для вывед-я продуктов
-усложн свзь кровеносн и выделит сист (формир-е сальп телец, затем нефронов)
Пол сист у позв связ с выд происхождение, положением и разделением канальцев.
Гомеостаз - это способность сохранять устойчивость и постоянство внутренней среды организма. За поддержание гомеостаза отвечают вегетативная нервная система и эндокринная система, которая контролируется гипоталамусом, а последний, в свою очередь, корой головного мозга.
Существует два механизма гомеостаза, отрицательная обратная связь и положительная обратная связь. Первая меняет реакцию системы на противоположную, вторая приводит к дестабилизирующему эффекту.
В качестве примера можно привести сохранение постоянной температуры тела, сохранение определенного числа особей внутри сообщества, повышение и понижение кровяного давления при недостатке кислорода. У животных защитные функции выполняют перья, мех, большая масса тела. Те, кто не способен поддерживать активность в холодное время, впадают в спячку. Экзотермные животные имеют приспособления, помогающие регулировать температуру тела благодаря внешним источникам тепла. Изотермия свойственна только так называемым гомойотермным, или теплокровным, животным. Волосяной покров и жировая подкожная клетчатка предохраняют тело от охлаждения, а от перегрева оно защищено благодаря испарению влаги через потовые железы.
У теплокровных животных дыхат сист участвует в процессах терморегуляции. Терморегуляция проявляется в форме взаимосочетания процессов теплообразования и теплоотдачи, регулируемых нервно-эндокринным путем. Терморегуляцию принято разделять на химическую и физическую. Химическая терморегуляция осуществляется путем изменения уровня теплообразования, т. е. усиления или ослабления интенсивности обмена веществ в клетках организма. Физическая терморегуляция осуществляется путем изменения интенсивности отдачи тепла. Наиболее интенсивное теплообразование в организме происходит в мышцах. Даже если человек лежит неподвижно, но с напряженной мускулатурой, окислительные нродес-сы, а вместе с тем и теплообразование повышаются на 10%. Небольшая двигательная активность ведет к увеличению теплообразования на 50—80%, а тяжелая мышечная работа — на 400—500%. В условиях холода теплообразование в мышцах увеличивается, даже если человек находится в неподвижном состоянии. Это обусловлено тем, что охлаждение поверхности тела, действуя на рецепторы, воспринимающие холодовое раздражение, рефлекторно возбуждает беспорядочные непроизвольные сокращения мышц, проявляющиеся в виде дрожи (озноб). При этом обменные процессы организма значительно усиливаются, увеличивается потребление кислорода и углеводов мышечной тканью, что и влечет за собой повышение теплообразования. Температура крови печеночной вены выше температуры крови печеночной артерии, что указывает на интенсивное теплообразование в этом органе. При охлаждении тела теплопродукция в печени возрастает. Освобождение энергии в организме совершается за счет окислительного распада белков, жиров и углеводов. Поэтому все механизмы, которые регулируют окислительные процессы, регулируют и теплообразование. Теплоотдача осуществляется путем теплоизлучения (радиационная теплоотдача), конвекции, т. е. движения и перемешивания нагреваемого телом воздуха, теплопроведе-ния, т. е. отдачи тепла веществам, непосредственно соприкасающимся с поверхностью тела, /И испарения воды с поверхности кожи и легких. У человека в обычных условиях потеря тепла путем теплопроведения имеет небольшое/значение, так как воздух и одежда являются плохими проводниками тепла. Радиация, испарение и конвекция протекают с различной интенсивностью в зависимости от температуры окружающей среды. У человека в состоянии покоя при температуре воздуха ' около 20 °С и суммарной теплоотдаче, равной 419 кДж (100 ккал) в час, радиация составляет 66%, испарение воды —19%, конвекция -15% общей потери тепла организмом.
Выделительная система ланцетника – нефридиального типа. По строению близка к выделительной системе некоторых кольчатых червей. Нефридии у ланцетников – короткие изогнутые трубки (до 90 пар), расположенные в области глотки. Большая часть нефридия лежит в целоме. Имеется ряд отверстий (нефростомы), окруженных булавовидными клетками (соленоцитами). Соленоцит имеет тонкий каналец с мерцательным волоском, обеспечивающим ток целомической жидкости. Выводящий конец нефридия открывается в атриальную полость. Продукты метаболизма поступают в полость целома из капилляров, пронизывающих стенки целома, а затем за счет движения мерцательных волосков соленоцитов выводятся в атриальную полость и через атриопор – наружу.
Органы выделения у взрослых рыб представлены парными туловищными почками, парными мочеточниками, мочевым пузырем и мочевым отверстием. Конечными продуктами азотного обмена являются: аммиак, мочевина, триметиламиноксид (это вещество придает мясу рыб специфический запах).
Структурно-функциональная единица почки - нефрон. Проксимальная (начальная) часть нефрона - мальпигиево тельце, сост из бокалообразной почечной капсулы (капсула Боумена–Шумлянского) и расположенного в ее полости клубочка капилляров. Капилляры отделены от полости почечной капсулы особыми клетками (подоцитами). Полость капсулы Боумена–Шумлянского переходит в извитой каналец (который у рыб еще не дифференцирован на проксимальный и дистальный отделы, разделенные изгибом – петлей Генле). Из капилляров почечного клубочка в полость капсулы фильтруется жидкость, содержащая воду, углеводы, аминокислоты и различные ионы - первичная моча. В проксимальном отделе извитого канальца происходит обратное всасывание органических веществ, в дистальном отделе – всасывание воды. В дистальном отделе извитого канальца формируется вторичная моча с повышенным содержанием вредных для организма веществ. Извитые канальцы впадают в парные мочеточники, которые впадают в мочевой пузырь. В мочевом пузыре происходит дополнительное всасывание воды.
У пресноводных рыб возникает проблема выведения избытка воды и сохранения минеральных солей. Морские рыбы, наоборот, вынуждены экономить воду и выводить избыток солей. В регуляции водно-солевого обмена у рыб значительную роль играют жабры и кишечник. У проходных рыб происходит перестройка водно-солевого обмена при переходе из моря в реки и наоборот.
Выделительная и половая система Амфибий тесно связаны между собой и образуют мочеполовую систему, типичную для анамний. Основой выделительной системы являются туловищные почки. Основой половой системы у самок являются парные яичники, а у самцов – парные семенники.
Выделительная система. Парные компактные туловищные почки расположены по бокам позвоночного столба в задней части полости тела. У переднего края каждой почки располагаются жировые тела – резерв питательных веществ для формирования половых клеток. Структурно-функциональной единицей почек является нефрон (его строение и функционирование принципиально не отличается от такового у рыб). Основным продуктом азотного обмена у лягушки является мочевина, а у головастиков и Хвостатых амфибий, живущих в воде – аммиак. По внешнему краю каждой почки тянется тонкий канал – мочеточник (вольфов канал), открывающийся в клоаку. В клоаку открывается и мочевой пузырь. Моча накапливается в мочевом пузыре, концентрируется а затем выводится наружу.
Date: 2015-09-24; view: 1512; Нарушение авторских прав