Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Зависимость активности RyR-канала от времени
|
|
В начале эксперимента по исследованию активности RyR-каналов все каналы в ансамбле были закрыты, а в момент времени t=0 при cis[Ca]>0 происходил быстрый процесс активации и дальнейшей процесс медленной адаптации каналов. Регистрация активности канала в численных экспериментах проводилась, после выхода на стационарный режим, начиная с момента t =3 c.
На рисунке 3.26 представлены результаты численных экспериментов по наблюдению активности RyR-канала при стационарных условиях. Данный эксперимент проводился при следующих значениях интенсивностей переходов: 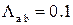 мс-1,
мс-1, 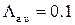 мс-1,
мс-1, 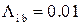 мс-1,
мс-1, 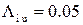 мс-1,
мс-1, 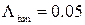 мс-1. Эти значения соответствуют значению cis[Ca]=0.5 мкМ согласно формуле (2.6). Активность канала исследовалась на малых интервалах длительностью 10 мс, что позволило построить график зависимости вероятности пребывания RyR-канала в открытом состоянии от времени (рис. 3.27б).
мс-1. Эти значения соответствуют значению cis[Ca]=0.5 мкМ согласно формуле (2.6). Активность канала исследовалась на малых интервалах длительностью 10 мс, что позволило построить график зависимости вероятности пребывания RyR-канала в открытом состоянии от времени (рис. 3.27б).
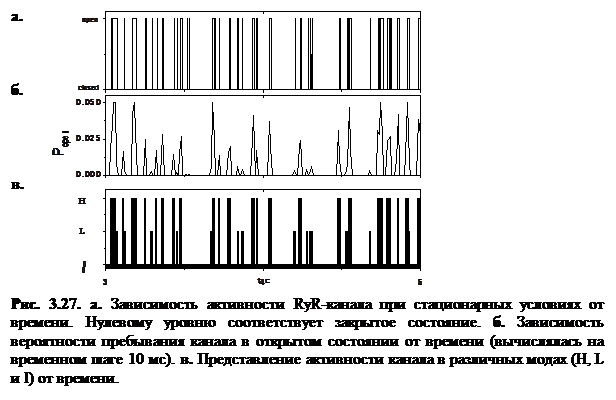
Как видно из графика, зависимость вероятности пребывания канала в открытом состоянии 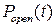 является неоднородной, и динамика RyR-канала при стационарных условиях носит стохастический характер.
является неоднородной, и динамика RyR-канала при стационарных условиях носит стохастический характер.
Гистограмма распределения вероятности пребывания канала в открытом состоянии (рис. 3.28) имеет два локальных максимума в точках, соответствующих 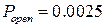 и
и  . Локальный минимум соответствует значению
. Локальный минимум соответствует значению 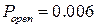 и определяет граничное значение вероятности пребывания канала в открытом состоянии
и определяет граничное значение вероятности пребывания канала в открытом состоянии 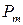 [121], которое разделяет так называемые моды активности RyR-канала на текущем интервале. При значении
[121], которое разделяет так называемые моды активности RyR-канала на текущем интервале. При значении 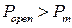 интервал характеризуется высокой активностью (мода Н), при
интервал характеризуется высокой активностью (мода Н), при 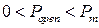 канал находится в моде низкой активности L. Значение вероятности
канал находится в моде низкой активности L. Значение вероятности 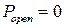 определяет нулевую активность канала (мода I). Зависимость положения канала в модах H, L и I в зависимости от времени изображена на рисунке 3.27в. Информация о положении канала в той или иной моде позволяет более детально описать его активность в текущий момент времени.
определяет нулевую активность канала (мода I). Зависимость положения канала в модах H, L и I в зависимости от времени изображена на рисунке 3.27в. Информация о положении канала в той или иной моде позволяет более детально описать его активность в текущий момент времени.
3.7.2 Зависимость активности RyR-канала от уровня cis[Ca]
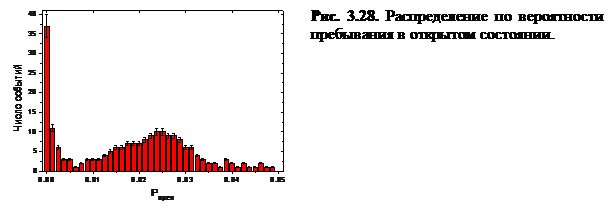
На рисунке 3.29 изображены зависимости активности RyR-канала от времени при различных значениях cis[Ca] (открытое состояние соответствует значению 1).
Для сравнения результатов численных реализаций с экспериментальными данными, на рисунке 3.30 представлены графики экспериментальные регистрации активности RyR-канала при различных значениях cis[Ca] [15].

Сравнивая рисунок 3.30а с результатами моделирования (рис. 3.30б), можно сделать вывод, что результаты численных экспериментов находятся в хорошем согласии с экспериментальными данными.
В работе при проведении анализа кинетических характеристик динамики RyR-канала в рамках модели, были построены графики зависимости 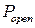 от концентрации Са2+ в cis-части при различных значениях числа кластеров в активных центрах канала, представленные на рисунке 3.31.
от концентрации Са2+ в cis-части при различных значениях числа кластеров в активных центрах канала, представленные на рисунке 3.31.
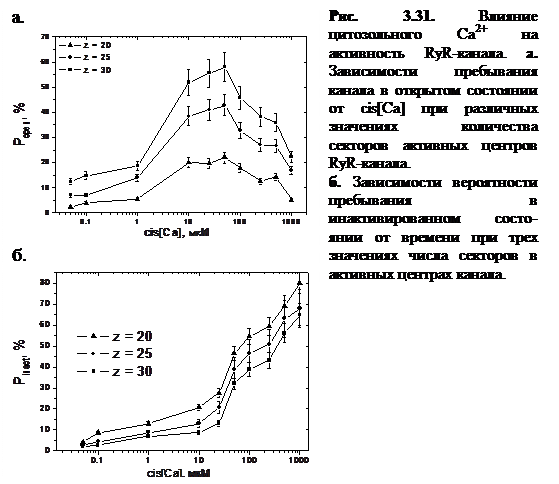
Зависимость активности RyR-канала при стационарных условиях от концентрации цитозольного Са2+ носит колоколообразный характер: 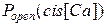 растет с увеличением cis[Ca] в интервале от 0 до 100 мкМ и спадает при cis[Ca]>100 мкМ.
растет с увеличением cis[Ca] в интервале от 0 до 100 мкМ и спадает при cis[Ca]>100 мкМ.
Эффект понижения 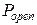 при достаточно больших значениях cis[Ca] получил в литературе название Са2+-зависимой инактивации [24] и является одним из ключевых феноменов тонкой регуляции RyR-канала ионами Са2+. Однако следует отметить, что концентрации cis[Ca]>100 мкМ не являются физиологическими и представляют интерес только для экспериментов in vitro.
при достаточно больших значениях cis[Ca] получил в литературе название Са2+-зависимой инактивации [24] и является одним из ключевых феноменов тонкой регуляции RyR-канала ионами Са2+. Однако следует отметить, что концентрации cis[Ca]>100 мкМ не являются физиологическими и представляют интерес только для экспериментов in vitro.
Приведенные на рисунке 3.31б графики зависимости вероятности пребывания в инактивационном состоянии (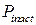 ) от cis[Ca] говорят о том, что с ростом cis[Ca] увеличивается частота электронных переходов в инактивационное состояние, в связи с этим повышается .
) от cis[Ca] говорят о том, что с ростом cis[Ca] увеличивается частота электронных переходов в инактивационное состояние, в связи с этим повышается .
При исследовании динамических рядов активности RyR-канала при различных значениях cis[Ca] были изучены моды активности канала и построена гистограмма распределения по пребываниям в той или иной моде, представленная на рисунке 3.32.
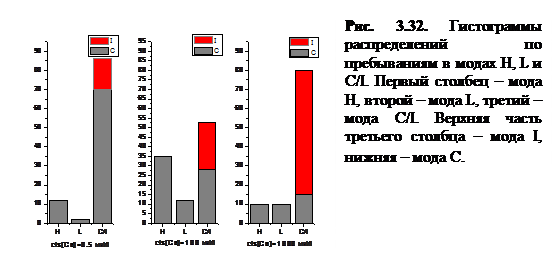
Были введены следующие моды активности:
· Мода С/I соответствует пребыванию канала в закрытом состоянии С или в инактивационном состоянии I. На гистограмме состояния С и I разделены в третьем столбце. Вероятность пребывания в открытом состоянии в этой моде является нулевой.
· Моде H соответствуют длительные пребывания в открытом состоянии О и частые переходы из закрытого состояния в открытое (при этом время пребывания в открытом состоянии больше, чем в закрытом: 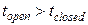 ). В данной моде .
). В данной моде .
· Моде L соответствует серия кратковременных пребываний в открытом состоянии (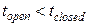 ). Закрытия происходят вследствие переходов в конформационно закрытое состояние и в инактивацинное состояние. В моде низкой активности: .
). Закрытия происходят вследствие переходов в конформационно закрытое состояние и в инактивацинное состояние. В моде низкой активности: .
Как ясно из рисунков 3.31 и 3.32, с ростом cis[Ca] наблюдается увеличение вероятности нахождения системы в инактивационном состоянии, и оно становится превалирующим после значения cis[Ca]=100 мкМ, которое соответствует максимуму зависимости .
Согласно экспериментальным данным [20], активность RyR-каналов в кардиомиоцитах становится нулевой при очень высоких значениях концентрации цитозольного Са2+ (5-10 мМ), которые, предположительно, недостижимы в клетках. Однако, Са2+-зависимая инактивация также как и адаптация, является механизмом негативного контроля, обеспечивающим своевременное завершение процесса высвобождения Са2+ из СР во время клеточного цикла.
3.7.3 Влияние ионов Mg2+ на динамику одиночного RyR-канала
Ионы Mg2+ как и ионы Са2+ обладают сильным сродством с активационной частью RyR-канала, однако, занимая вакантные места на активационных частях, они не приводят к открытию канала [122, 123].
Как и в случае взаимодействия ионов Са2+с активными центрами, в настоящей диссертационной работе предлагается статистический подход к рассмотрению конкурентного присоединения ионов Mg2+ и Са2+ к активным центрам канала.
На рисунке 3.33а схематично представлен процесс присоединения ионов Са2+ и Mg2+ с активному центру канала. Заполнение ионами активных мест связывания активного центра описано с помощью диаграммы Эйлера (рис. 2.31б).
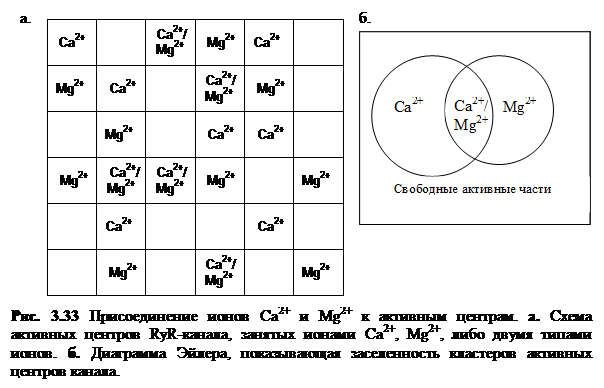
Было сделано предположение, что если в кластер активного центра попадают оба типа ионов, то вероятность присоединения того или иного типа ионов к свободному центру равняется ½.
Пусть 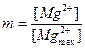 – относительная концентрация ионов Mg2+ вблизи канала, где
– относительная концентрация ионов Mg2+ вблизи канала, где 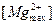 – максимальная концентрация магния, при которой заполнены все места связывания активного центра канала;
– максимальная концентрация магния, при которой заполнены все места связывания активного центра канала;  – относительная концентрация ионов Са2+.
– относительная концентрация ионов Са2+.
Далее определялась вероятность нахождения более чем k ионов Са2+ в активном центре канала с учётом присутствия на нем ионов Mg2+.
Относительное число активных мест, в которых находятся только ионы Са2+ в кластере: 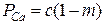 ; относительное число секторов, в которых находятся ионы Са и Mg, но присоединяются ионы Са:
; относительное число секторов, в которых находятся ионы Са и Mg, но присоединяются ионы Са: 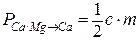 . Соответственно, относительное число «сработавших» ионов Са2+ равно
. Соответственно, относительное число «сработавших» ионов Са2+ равно 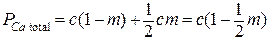 . Число комбинаций k ионов Са2+ в z частях кластера равно
. Число комбинаций k ионов Са2+ в z частях кластера равно 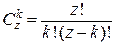 . Наконец, вероятность обнаружения более чем k ионов Са2+ в кластере активной части равняется:
. Наконец, вероятность обнаружения более чем k ионов Са2+ в кластере активной части равняется:
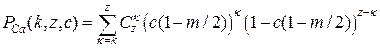 . (3.21)
. (3.21)
В численных экспериментах значение cis[Ca] варьировалось в диапазоне от 0.05 до 1000 мкМ. В первой части экспериментов не учитывалось влияние ионов Mg2+, и интенсивность электронных переходов в зависимости от cis[Ca] рассчитывалась по формуле 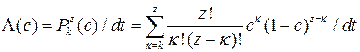 (см. формулу 2.10), где с – относительная концентрация ионов Са2+ в cis-части.
(см. формулу 2.10), где с – относительная концентрация ионов Са2+ в cis-части.
Во второй серии экспериментов учитывалось присутствие Mg2+ в растворе (Mg=3 мМ), и 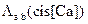 и
и 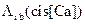 рассчитывались согласно формуле (3.21). Графики зависимостей интенсивностей электронных переходов с учетом и без учета Mg2+ представлены на рисунке 3.33.
рассчитывались согласно формуле (3.21). Графики зависимостей интенсивностей электронных переходов с учетом и без учета Mg2+ представлены на рисунке 3.33.
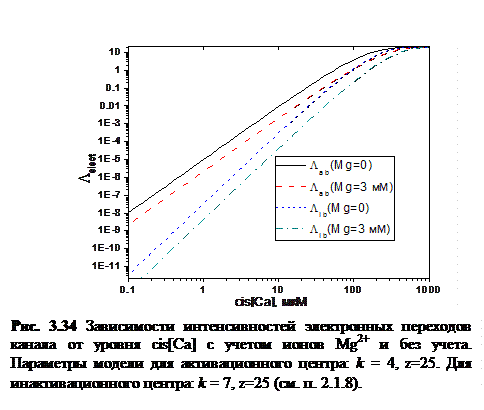
Как видно из рисунка, учет Mg2+ сдвигает зависимость 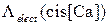 вниз, то есть статистический подход хорошо описывает ингибирующие свойства ионов Mg2+.
вниз, то есть статистический подход хорошо описывает ингибирующие свойства ионов Mg2+.
На рисунке 3.35 представлены графики зависимости вероятности пребывания канала в открытом состоянии от концентрации Са2+ в cis-части в отсутствии и в присутствии ионов Mg2+ 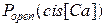 .
.

Как видно из графика, зависимости имеют колоколообразную форму с максимумом, соответствующим значению  cis[Ca]=50 мкМ (без учета Mg2+) и cis[Ca]=100 мкМ (с учетом Mg2+). Другими словами, при учете ионов Mg2+ происходит сдвиг вправо зависимости
cis[Ca]=50 мкМ (без учета Mg2+) и cis[Ca]=100 мкМ (с учетом Mg2+). Другими словами, при учете ионов Mg2+ происходит сдвиг вправо зависимости 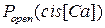 . Для сравнения на рисунке 3.35б приведен график из работы [20]. Анализ представленных кривых говорит о том, что результаты моделирования с высокой степенью точности совпадают с экспериментальными данными.
. Для сравнения на рисунке 3.35б приведен график из работы [20]. Анализ представленных кривых говорит о том, что результаты моделирования с высокой степенью точности совпадают с экспериментальными данными.
В присутствии ионов Mg2+ в растворе, в котором находится RyR-канал, график зависимости сдвигается вниз и вправо, то есть максимум достигается при больших значениях 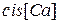 и максимум функции меньше, чем в отсутствие ионов Mg2+, конкурирующих с активными ионами Са2+.
и максимум функции меньше, чем в отсутствие ионов Mg2+, конкурирующих с активными ионами Са2+.
Ионы Mg2+ являются потенциальными ингибиторами RyR-каналов, в связи с этим находят активное применение в фармакологии и терапии для понижения частоты высвобождений Са2+ для предотвращения аритмии и других патологий.
Модификация электронно-конформационной модели и введение инактивационного состояния позволили описать не только основные свойства изолированных RyR-каналов, но и важнейшие тонкие эффекты взаимодействия каналов с ионами Са2+, которые являются механизмами негативного контроля и должны учитываться при моделировании процесса КВВК.
Заключение
В данной главе приведены результаты параметрического анализа электронно-конформационной модели одиночного RyR-канала, сравнения результатов ЭК-модели с традиционными теориями, основанными на марковских цепях.
В рамках ЭК-модели проведена серия численных экспериментов по исследованию:
· процессов открытия RyR-канала при резком повышении уровня cis[Ca];
· процесса закрытия RyR-канала при резком уменьшении уровня cis[Ca];
· процесса адаптации RyR-канала к установившемуся уровню cis[Ca];
· зависимости вероятности пребывания RyR-канала в открытом и закрытом состояниях при постоянном уровне cis[Ca];
· влияния ионов Mg2+ на активность RyR-канала при стационарных условиях.
· зависимости конформационной координаты Q от времени методом Херста.
Сравнительный анализ показал, что результаты проведенных численных экспериментов хорошо согласуются с известными экспериментальными данными, и ЭК-модель дает адекватное описание стохастической динамики RyR-канала.
ГЛАВА 4. Моделирование динамики ионов Са2+ между отделами кардиомиоцита
В данной главе диссертационной работы описаны эффекты, связанные с кооперативной динамикой RyR-каналов в кластере высвобождающей единицы. Также представлены результаты параметрического анализа модели динамики ионов Са2+ между отделами клетки водителя сердечного ритма.
Date: 2015-08-24; view: 602; Нарушение авторских прав