Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
I – инспираторные (вдох). 3 page
|
|
находится в опасности, то гормоны мозгового вещества создают все условия для успешной борьбы или не менее успешного бегства. Многообразие эффектов адреналина и норадреналина связано с тем, что в различных тканях они реагируют с имеющимися там несколькими видами рецепторов.
Патологии мозгового вещества надпочечников возникают редко, однако при развитии в этой области опухоли (феохромоцитомы) происходят приступообразные выбросы адреналина, причем симптомы, наблюдаемые при этом, те же, что и при избыточном возбуждении симпатического отдела вегетативной нервной системы. Клинически феохромоцитома проявляется подъемом артериального давления (злокачественная гипертензия), не поддающаяся медикаментозному лечению.
Единственный метод лечения — хирургический. Любое разрушение или искусственное удаление надпочечников (адреналэктомия) приводит к быстрой гибели. Эффекты гормонов мозгового слоя надпочечников очень близки к эффектам, возникающим при возбуждении симпатического отдела вегетативной нервной системы, и обе эти системы, каждая своими путями, обеспечивает выживание организма в неблагоприятных условиях существования.
Кора надпочечников вырабатывает кортикостероиды. Часть из них («минералокортикоиды») регулирует обмен калия и натрия в почках. Вторая часть («глюкокортикоиды», ос-новной представитель кортизол) управляет обменом глюкозы в организме, в частности, усиливает ее образование из других веществ (белков, жиров); особенно ярок этот эффект при стрессе. В целом глюкокортикоиды перестраи-вают обмен веществ на усиленное потребление энергии, «сопровождая» эффекты симпатической НС и адреналина. Глюкокортикоиды, кроме того, тормозят активность иммунной системы и развитие воспаления, что позволяет использовать их как противовоспалительные препараты.
Выделение глюкокортикоидов усиливает кортикотропин = адренокортикотропный гормон (АКТГ или АСТН). Выделение АКТГ активирует кортиколиберин (CRH). Кортизол тормозит выброс АКТГ и CRH.
АКТГ (АСТН) состоит из 39 аминокис-лот. Его фрагменты длиной 7-10 а/к способны проникать в ЦНС, оказывая ноотропное действие, улучшая обуче-ние и память, снижая проявления депрессивности (препарат СЕМАКС). Кортиколиберин (CRH) стимулирует двигательную активность и эмоциональные проявления; снижает пищевую и половую мотивации. В последнее время система CRH - АКТГ – глюкокортикоиды интенсивно исследуется в связи с возможностями управления весом организма.
7-2. Синтез, взаимопревращения и инактивация глутаминовой кислоты и ГАМК. Значение аминокислот для деятельности мозга.

Медиаторы-аминокислоты являются главными медиаторами ЦНС.
Глутаминовая кислота (Glu) – главный возбуждающий меди-атор (около 40% всех нейронов); проведение основных потоков информации в ЦНС (сенсорные сигналы, двигат. команды, память).
Гамма-аминомасляная кислота (ГАМК) – главный тормозный медиатор (также около 40% всех нейронов); запрет проведения «ненужной» информации (внимание, двигательный контроль).
Глицин – вспомогательный тормозный медиатор (менее 1%; основная функция – торможение мотонейронов).
Нормальная деятельность ЦНС обеспечивается тонким балансом Glu и ГАМК.
При нарушении баланса Glu и ГАМК:
· синдром дефицита внимания и гиперактивности детей (СДВГ);
· повышенная нервозность и тревожность взрослых;
· нарушения сна, бессонница;
· эпилепсия (чаcто врожденная патология, у 0.5% населения).
В клинических целях используют агонисты ГАМК и антагонисты Glu. Антагонисты ГАМК и агонисты Glu – яды, вызывающие судороги (конвульсанты).
Глутаминовая кислота. Пищевая аминокислота – входит в состав белков пищи и белков нашего тела; самая распространенная: потребляем с едой 5-10 г в сутки.Стандартная структура; радикал -СН 2 -СН 2 -СООН.В водных растворах существует в ионизированном виде, то есть в форме отрицательно заряженного остатка глутамата («глютамата»).
Для синтеза необходимы: a-кетоглутаровая кислота (промежуточный продукт окисления глюкозы в цикле Кребса; в больших количествах образуется в митохондриях); аминогруппа любой а/к, полученной с пищей; фермент из группы аминотрансфераз. Такой синтез идет во всех клетках организма.
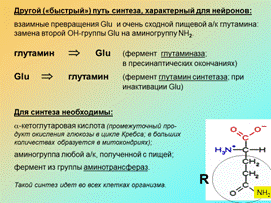
Другой («быстрый») путь синтеза, характерный для нейронов: взаимные превращения Glu и очень сходной пищевой а/к глутамина: замена второй ОН-группы Glu на аминогруппу NH 2.
глутамин Þ Glu (фермент глутамин синтетаза;
в пресинаптических окончаниях)
Glu Þ глутамин (фермент глутаминаза; при
инактивации Glu)
Для синтеза необходимы: a-кетоглутаровая кислота (промежуточный продукт окисления глюкозы в цикле Кребса; в больших количествах образуется в митохондриях);
аминогруппа любой а/к, полученной с пищей; фермент из группы аминотрансфераз. Такой синтез идет во всех клетках организма.
Во всех тканях организма (в т.ч. в мозге) очень много Glu. В связи с этим долгое время не могли поверить, что столь распространенное вещество является медиатором ЦНС. Однако это именно так. Дело в том, что пищевой глутамат почти не преодолевает ГЭБ, и для выпол-нения медиаторных функций Glu синтезируется непосредственно в пресинаптических окончениях из глутамина; определенный вклад вносит также образование Glu из a-кетоглутаровой кислоты (a-KG).
После синтеза Glu загружается в везикулы (◄), выбрасывается в синаптическую щель при приходе ПД и влияет на рецепторы (↓↓↓), запуская ВПСП.
Введение Glu непосредственно в мозг (в желудочки) вызывает возбуждение ЦНС и судороги.
Сходные эффекты наблюдаются при отравлении агонистами Glu, часть из которых является токсинами растений. Пример: домоевая кислота; вырабатывается некоторыми одноклеточными водорослями; токсин накапливается в телах животных, поедающих фитопланктон (двустворчатые моллюски, некоторые рыбы) и способен отравлять птиц, млекопитающих, человека. Смертельные случаи: западное побережье Канады; «бешенство» птиц в Калифорнии (Дюморье, Хичкок).
Инактивация Glu: Из синаптический щели Glu l переносится в глиальные клетки, где превращается в глутамин (Gln) l (с помощью фермента глутаминазы). Глутамин затем может перемещаться в пресинаптическое окончание и вновь становиться Glu в митохондриях.Часть Glu возвращается из синаптической щели прямо в пресинаптич. окончание («обратное всасывание»).
ГАМК. Непищевая аминокислота:аминогруппа в 3-м (g) положении.Синтез – из глутаминовой кислоты за счет отщепления СО2(фермент глутамат декарбоксилаза).
ГАМК может использоваться в качестве медиатора, но может терять аминогруппу (фермент ГАМК-трансфераза) и быстро окисляться с выдел. энергии.
Именно для нейронов характерна следующая цепь реакций: глюкоза ® … ® a-кетоглутаровая кислота ® Glu ® ГАМК ® … ® энергия
ГАМК на 10% проходит ГЭБ. Однако при ее системном введении (таблетки «Аминалон») наблюдается не столько торможение ЦНС, сколько «питание» нейронов и улучшение их общего состояния (ноотропное действие ГАМК).
Инактивация ГАМК. В значительной мере сходна с инактивацией Glu: обратное всасывание ГАМК в пресинаптическое окончание либо всасывание глиальными клетками. В последнем случае ГАМК разрушается ферментом ГАМК трансферазой: аминогруппа ГАМК переносится на a-кетоглутаровую кислоту с образованием Glu. Далее Glu может превращаться в глутамин, который транспортируется в пресинаптическое окончание, опять становится Glu, а из Glu образуется ГАМК. Круг замкнулся.
Инактивация ГАМК происходит в основном путем обратного всасывания в пресинаптическое окончание и последующего превращения в глутаминовую кислоту. Реакцию эту осуществляет особый фермент ГАМК-трансфераза. Его блокаторами являются соли вальпроевой кислоты. Они оказывают транквилизирующее и противосудорожное действие. Вальпроаты особенно полезны при малых формах эпилепсии. Вызываемые ими побочные депрессантные изменения, как правило, минимальны.
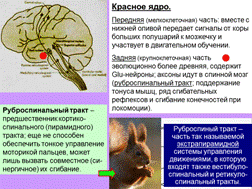
7-3. Средний мозг: четверохолмие и реакция на новизну. Управление движениями глаз. Функции различных областей красного ядра и черной субстанции.
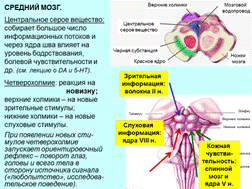

Движения глаз:
с каждым глазом связано по 6 мышц, управляемых III, IV и VI нервами;
два основных типа движений глаз – слежения и саккады (быстрые скачки);
в основе врожденные программы, но мы учимся ими управлять (вначале – произвольная коррекция, а затем – автоматизация);
тесты на рассматривание картинок – еще одно «окно в бессознательное».
(Чтение: [1] – скачок в начало строки; [2] мини-саккады (5-7 скачков вдоль строки, текст читается «в несколько приемов»).)

8-1. Щитовидная железа и эпифиз: выделяемые гормоны и их эффекты (в т.ч. действие на нервную систему). ЦНС и управление работой щитовидной железы и эпифиза.
Щитовидная железа: выделяет йод-содержащие гормоны тироксины, усиливающие обмен веществ (образование энергии) во всех клетках организма, в т.ч. в мозге
Выделение тироксинов (Т4 и Т3) усиливает тирео-тропный гормон передней доли гипофиза (TSH; тиреостимулирующий гормон). Гипоталамус, измеряя концентрацию тироксинов и TSH в крови, усиливает выделение статина (его роль играет дофамин) либо либерина (тиролиберина, TRH; является также либерином пролактина).
Тиролиберин активирует выброс TSH.
Опасен как дефицит, так и избыток тироксинов в организме.
При дефиците (например, из-за нехватки йода в пище) – снижение обмена веществ, вялось, депрессии («микседема»); у новорожденных – оставание умственного развития.
При избытке – нервозность, бессонница, повышенный аппетит и худоба, гиперактивность симпатичес-кой НС, «выпученные» глаза.
Причиной в обоих случаях могут быть аутоиммунные нарушения.
Влияние гормонов щитовидной железы (из учебника): программирующие гормоны (пр.участвуют в половом созревании), усиливают обмен веществ, стимулируя клеточное дыхание, повышают секрецию соматотропного гормона гипофиза, усиливают производство тепла организмом-термогенез
Гипофункция щитовидной железы-кретинизм (детство), микседема (взрослый)
Гиперфункция-базедова болезнь

Мелатонин – гормон эпифиза.
Синтезируется из триптофана (как и серотонин). «Гормон сонного состояния»: снижает активность обмена веществ во многих внутренних органах и ЦНС. Выделением управляют центры сна и бодрствования
На свету паравентрикулярные ядра гипоталамуса (PVN) через нейроны бокового рога серого вещества спинного мозга (ILC) и шейные симпатические ганглии (SCG) сдерживают выделение мелатонина. В темноте супра-хиазменные ядра (SCN) акти-вируют выделение мелатонина за счет того, что тормозят PVN с помощью GABA (т.е. ГАМК).
Мелатонин используется в
качестве лекарства при бессон-нице (облегчает засыпание)
Эпифиз — маленькая железа, массой всего 170 мг, являющаяся выростом крыши третьего желудочка мозга и расположенная между верхними буграми четверохолмия. Гормон эпифиза — мелатонин по своему химическому строению является модификацией медиатора ЦНС серотонина, который в свою очередь синтезируется из аминокислоты
триптофана. Секреция мелатонина уменьшается на свету и усиливается в темноте. Показано увеличение секреции этого гормона у слепых людей. Мелатонин обладает мембранной рецепцией и действует через систему G-белков на активность аденилатциклазы, подавляя ее. Под воздействием мелатонина происходят изменения в обмене черного пигмента — меланина, локализованного в коже, радужке, сетчатке, некоторых структурах мозга, в результате кожа светлеет, повышается чувствительность фоторецепторов сетчатки. Мелатонин участвует в обеспечении суточных ритмов человека, в том числе и ритмов секреции ряда гормонов. Мелатонин —
антигонадотропный гормон. Его секреция снижается при половом созревании, что стимулирует замыкание системы гипоталамус — гипофиз — гонады. Мелатонин обладает целым рядом воздействий на функции мозга. Так, увеличенное образование и секреция его в темноте может приводить к депрессии у лиц, вынужденных бодрствовать и работать в темное время суток. Повышенный уровень мелатонина индуцирует сонливость, вялость, может вызвать длительный глубокий сон. По-видимому, угнетая синтез некоторых факторов, усиливающих рост тканей, ускоряющих деление клеток, мелатонин может тормозить развитие опухолей.
8-2. Два типа рецепторов ацетилхолина (Ацх), их свойства. Агонисты и антагонисты рецепторов Ацх: примеры веществ и разнообразие эффектов.
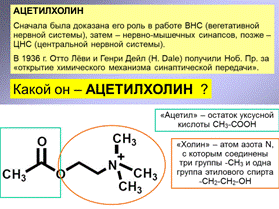

Функции Ацх-рецепторов(всех)
Их функцию можно определить, как нормализующую тонус мозга (т.е. при утомлении активируют ЦНС, при перевозбуждении – успокаивают).



Агонисты, Антагонисты



Блокаторы (на всякий случай)
Примеры блокаторов:
токсин малабарских бобов эзерин (физостигмин);
фосфорорганические инсекти-циды (хлорофос, дихлофос и т.п.; могут вызывать токсикоманию);
боевые нервно-паралитические газы (зарин, табун).
Пиридостигмин и сходные препараты, а также ряд агонистов рецепторов Ацх используются для лечения болезни Альцгеймера – самого распространенного нейродегенеративного заболевания, при котором первыми страдают Ацх-нейроны больших полушарий.
Прозерин и более мягко действующий пиридостигмин, соз-данные на основе эзерина, исполь-зуют при миастении (аутоим-мунное заболевание: антитела атакуют никотиновые ре-цепторы; развивается мышечная слабость, вялость, быстрая утомляемость; характерн. признак – опущенные веки). Основное лечение – иммуносупрессия
8-3. Первичная слуховая кора и тонотопия. Вторичная и третичная слуховая кора: опознавание слуховых образов разной степени сложности.
Слуховая кора:
Первичная – височная доля, по границе боковой борозды.
Завершение частотно-амплитудного анализа, наиболее точная тонотопическая карта.
В передних зонах – низкие частоты; особенно детально анализируется речевой диапазон – 50-500Гц
Ниже расположена вторичная слуховая кора – опознавание звуковых образов как совокупности частот (шумы, «звуки природы» и т.п.).
Как правило, свойства нейронов этой области – результат обучения.
Невербальная коммуникация (плач, смех и т.п.) опознается врожденно.
Задняя часть височной доли – третичная слуховая кора: узнавание наиболее сложных слуховых образов (музыки, речи). Узнавание речи на слух - зона Вернике)
Основная проблема: нужно реагировать не на частоты и их совокупность, а общую форму спектра (вне зависимости от тональности).
Зона Брока – речедвигательный центр.
Тонотопическая карта:
Улитка – частотно-амплитудный анализатор («на выходе» возникает спектр звука). В ЦНС – тонотопические карты.
Завершение частотно-амплитудного анализа, наиболее точная тонотопическая карта.
9-1. Половые железы; половые гормоны и их эффекты (в т.ч. действие на нервную систему). ЦНС и управление работой половых желез.
Кроме того, передн. доля гипофиза вырабатывает тропные гормоны:
тиреотропный (тиреостимулирующий – TSН; влияет на щитовидную железу);
адренокортикотропный (АСТН; влияет на кору надпочечников);
гонадотропные FSH и LH влияют на половые железы мужчин и женщин;
соматотропный (гормон роста) – на рост тела, его общий размер
Гипоталамус: Статины и либерины выделяются в кровь нейроэндокринными клетками серого бугра, измеряющими содержа-ние в крови «конечного» гормона (ти-роксина, половых гормонов и др.).
Два следующих тропных гормона(гипофиз) регулируют работу половых желез:
LH и FSH – лютеинизирующий и фолликулостимулирующий гормоны («гонадотропины»).
Несмотря на названия, в равной мере влияют на семенники мужчин и яичники женщин. Их функции:
активация синтеза и выделения половых гормонов;
стимуляция образования и созрева-ния половых клеток – сперматозои-дов и яйцеклеток (в пузырьках-фол-ликулах, далее следует овуляция).
Рилизинг-фактор гонадотропинов – пептид люлиберин (GnRH).
В ЦНС он активирует половое поведение, повышает эмоциональность, улучшает обучение.
Главными мужскими половыми гормонами являются тестостероны, тормозящие (ограничивающие) выделение люлиберина, LН и FSH.
Тестостероны активируют спермато-генез, у эмбриона – направляют развитие половой системы по мужскому типу; позже – определяют формирование мужских вторичных половых признаков.
(Вторичные половые признаки: борода, склонность к облысению, усиленный рост волос на теле, низкий голос, более мощное развитие мышц, склонность к отложению запасов жира в области живота, слабое развитие молочных желез.)
В ЦНС тестостероны влияют на половое влечение (либидо), половое поведение, агрессивность.
Главными женскими половыми гормонами являются прогестерон и эстрогены.
Прогестерон обеспечивает состояние готовности матки к беременности (содержание в крови максимально между овуляцией и менструацией).
Эстрогены «подталкивают» созрева-ние яйцеклеток, усиливают сокраще-ния яйцеводов и др. Они же опреде-ляют формирование женских вторич-ных половых признаков.(Вторичные половые признаки:
высокий голос, отсутствие бороды, склонность к отложению запасов жира в области бедер, развитие молочных желез и др)
В ЦНС эстрогены влияют на половое влечение и половое поведение.
9-2. Синтез глутаминовой кислоты (ГлК) в нервных клетках. Разнообразие рецепторов к ГлК; NMDA-рецепторы. Примеры и значение агонистов и антагонистов ГлК.
Глутаминовая кислота (Glu) – главный возбуждающий меди-атор (около 40% всех нейронов); проведение основных потоков информации в ЦНС (сенсорные сигналы, двигат. команды, память).
Глутаминовая кислота.
Пищевая аминокислота – входит в состав белков пищи и белков нашего тела; самая распростра-
ненная: потребляем с едой 5-10 г в сутки.
Стандартная структура; радикал -СН2-СН2-СООН.
В водных растворах существует в ионизированном виде, то есть в форме отрицательно заряженного остатка глутамата («глютамата»).




Агонисты Glu:
Введение Glu непосредственно в мозг (в желудочки) вызывает возбуждение ЦНС и судороги.
Сходные эффекты наблюдаются при отравлении агонистами Glu, часть из которых является токсинами растений. Пример: домоевая кислота; вырабатывается некоторыми одноклеточными водорослями; токсин накапливается в телах животных, поедающих фитопланктон (двустворчатые моллюски, некоторые рыбы) и способен отравлять птиц, млекопитающих, человека. Смертельные случаи: западное побережье Канады; «бешенство» птиц в Калифорнии).
Глутамат, помимо действия на рецепторы постсинаптической мембраны, способен влиять на вкусовые клетки-рецепторы языка («вкус белка»). Существуют особые клетки-рецепторы для сладкого, горького, кислого, солёного и глутамата. На мембране – белки-рецепторы к соответствующим веществам. Их активация ведёт к входу Ca2+, выбросу Glu (как медиатора) и возникновению ПД в волокнах вкусовых нервов (VII и IX).
Umami – яп. «мясной»; термин для описания особого «бульонного» вкуса морской капусты, соевого соуса, сыров (пармезан), грибов и т.п. В начале XXв. было показано, что это – вкус глутамата. С тех пор глутамат и его производные всё шире используются как «усилители вкуса» (E620 и др.). Избыток Glu (10г и более одномоментно) может вести к головной боли, потоотделению, сердцебиению («синдром китайского ресторана», не путать с пищевой аллергией).
9-3. Дыхательный центр продолговатого мозга и моста, принципы его функционирования. Роль нейронов-пейсмекеров, механорецепторов легких, хеморецепторов.

Среди нейронов вдоха ключевую роль играют клетки-пейсмекеры, находящиеся в ядрах нижней части ромбовидной ямки.
Врожденно обусловленная частота их активации у человека: примерно 1 волна в 5 сек (12 раз в мин = частота дыхания во сне
От клеток-пейсмекеров (генераторов ритма) ПД передаются к другим дыхат. нейронам и мотонейронам шейных и грудных сегментов спинного мозга, запускающим сокращение диафрагмы и межреберных мышц.

Еще о дыхательных центрах:
инспираторные нейроны – это не только пейсмекеры, но и клетки, «зацикливающие» ПД по замкнутому контуру, что дает возможность оказывать на мотонейроны стабильное активирующее действие;
хеморецепторы СО2 (и Н+) представляют собой нейроны на дне ромбовидной ямки; активируются в основном при физической нагрузке;
хеморецепторы О2 расположены в каротидном синусе (область разветвления на наружную и внутреннюю сонные артерии); важны, например, при подъеме в горы (на высоте 5 км воздуха в 2 раза меньше);
пробуждение приводит к активации пейсмекеров центрами бодрствования, и частота дыхания возрастает до 16-18/мин; при эмоциях и физической нагрузке – до 30-40/мин.
Передача инфор-мации о содержа-нии О2 в крови идет по волокнам IX нерва (кроме того, на схеме показана область, где распо-ложены рецепторы растяжения аорты; сигнал идет по волокнам Х нерва).


10-1. Потенциал покоя (ПП) нервных клеток. Роль Na+-K+-насоса и постоянно открытых ионных каналов. Уравнение Нернста, связь ПП с диффузией ионов Na+ и К+.
Сигнал по мембране нейрона передаётся в виде коротких электрических импульсов – потенциалов действия (ПД). Этот процесс можно сравнить с передачей информации с помощью включения и выключения фонарика (ПД= «вспышка света»).
Но для того, чтобы фонарик работал, нужна батарейка – источник электрической энергии. В случае нейрона таким источником является постоянный внутриклеточный заряд – потенциал покоя (ПП).
•Потенциал покоя нейрона – его постоянный отрицательный заряд, равный в среднем –70 мВ. Измерить ПП можно с помощью тончайшей, особым образом вытянутой стеклянной трубочки-микроэлектрода. Его кончик имеет диаметр <1 мкм, что позволяет практически без повреждения мембрану клетки. Микроэлектрод (в т.ч. канал внутри кончика) заполнен раствором соли, проводящим электрический ток. Это позволяет оценить, сравнить заряд цитоплазмы нейрона с зарядом межклеточной среды.
•Наличие ПП – результат жизнедеятельности нейрона, совместного функционирования всех биополимеров и органоидов клетки; погибший нейрон быстро теряет ПП. Первопричина ПП – разность концентраций ионов K+ и Na+ внутри и снаружи нейрона. Эту разность создаёт работа особого белка-насоса Na+-K+-АТФазы (Na+-K+-насоса).
•Na+-K+-АТФаза обменивает находящиеся внутри клетки ионы Na+ на захваченные в межклеточной среде ионы K+, затрачивая значительное количество АТФ.
В результате работы Na+-K+-АТФазы в нейроне оказывается примерно в 10 раз меньше Na+ и в 30 раз больше K+, чем в межклеточной среде.
• K+ “out”: K+ “in” = 1: 30
• Na+ “out”: Na+ “in” = 10: 1
Несмотря на всё это, до момента созревания (происходит на 2-3 месяце эмбрионального развития) нейрон не имеет заряда, и количество положительных (прежде всего, K+) и отрицательных ионов в его цитоплазме примерно одинаково. Признак созревания – появление на мембране нейрона постоянно открытых K+-каналов (определяется включением соответствующего гена). В результате становится возможной диффузия K+ из клетки.
• Как долго идёт диффузия K+ из нейрона?
Очевидный вариант («до выравнивания концентраций») неверен, поскольку двигаются заряженные частицы, и выход K+ сопровождается накоплением в цитоплазме отрицательного заряда. Этот отрицательный заряд мешает диффузии и в конце концов останавливает её. Возникает состояние «динамического равновесия»: число ионов K+, покинувших клетку благодаря диффузии = числу ионов K+, втянутых в клетку отрицательным зарядом цитоплазмы.
ПП – это отрицательный заряд цитоплазмы, останавливающий диффузию ионов K+ в межклеточную среду.
•«Уравнение Нернста»: ПП ~ lg (K+ “out”/K+ “in”)
Коэффициент пропорциональности равен 61.5 мВ для Т=36.6С; логарифм равен –1.48 (для соотношения концентраций 1/30).
С учётом этого ПП = -91 мВ («равновесный потенциал» для K+)
Такой вход Na+ ведёт к сдвигу заряда цитоплазмы вверх и частичной потере ПП (отсюда название – «ток утечки Na+»).
В реальной клетке ПП находится ближе к нулю (в среднем –70мВ). Причина: существование небольшого количества постоянно открытых каналов для ионов Na+. Избыток ионов Na+ в межклеточной среде, а также их притяжение к отрицательно заряженной цитоплазме приводят к входу Na+ в клетку.
•Ограничивает вход Na+,
• во-первых, малое число постоянно открытых Na+-каналов;
• во-вторых, работа Na+-K+-АТФазы, которая «откачивает» Na+, обменивая его на K+.
•В целом ПП зависит от 3х главных факторов:
• диффузии K+ из клетки;
• диффузии Na+ в клетку;
• работы Na+-K+-АТФазы.
Диффузия K+ из клетки определяется разностью концентраций K+ “out” и K+ “in”.
Если увеличить K+ “out”, то разность концентраций станет меньше, диффузия – слабее, и для её остановки потребуется не столь значительный ПП (произойдёт сдвиг заряда цитоплазмы вверх до достижения новой точки равновесия).
Если снизить K+ “out”, то разность концентраций станет больше, диффузия – сильнее, и для её остановки потребуется более значительный ПП (сдвиг заряда цитоплазмы).
Диффузия Na+ в клетку зависит, прежде всего, от концентрации постоянно открытых Na+-каналов на мембране. Эта величина, в свою очередь, является стабильным свойством конкретного нейрона. Чем больше таких каналов, тем ПП ближе к нулю, чем меньше – тем ПП ближе к уровню –91мВ.
Чем ближе ПП к нулю, тем возбудимее нейрон (такие нужны, например, в центрах бодрствования); чем ближе ПП к уровню –91 мВ, тем ниже возбудимость (минимальна в центрах, запускающих движения).
•Работа Na+-K+-АТФазы может быть нарушена химическими веществами, например, токсином одной из тропических лиан строфантином.
В этом случае ток утечки Na+ не будет полностью компенсироваться и ПП сместится в сторону нуля (степень смещения зависит от дозы токсина = доля заблокированных насосов).
Большая доза токсина настолько нарушает работу Na+-K+-АТФаз, что ПП теряется (происходит разрядка батарейки «фонарика»).
• Аналогия: Na+-K+-АТФаза -- «зарядное устройство» нейрона.
• Заключительная аналогия: лодка на поверхности водоёма.
Уровень воды = нулевой уровень; уровень бортов над водой= ПП (зависит от «веса лодки» = разность концентраций K+ во внешней среде и цитоплазме).
Ток утечки Na+ = отверстия в лодке, через которые втекает вода и снижает абсолютное значение ПП (приближая его к нулю).
Na+-K+-АТФаза – ковш, которым вычерпываем воду, удерживая лодку на плаву («поломка ковша» строфантином приведёт к тому, что лодка утонет)
Date: 2016-11-17; view: 502; Нарушение авторских прав