Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Минимальные наборы генов, замещение неортологичных генов (ЗНОГ) и ускользающее незаменимое ядро жизни
|
|
Секвенирование геномов симбиотических и паразитических бактерий привело к соблазнительной идее, что их генетический репертуар может быть близок к «наименьшему возможному набору генов», то есть такому, который является необходимым и достаточным для поддержания простой (прокариотической) клетки при самых благоприятных условиях, какие только могут существовать вне других клеток (Fraser et al., 1995; Mushegian and Koonin, 1996b). Последнее условие чрезвычайно важно, поскольку «наименьшим возможным» набор генов будет лишь в отношении к окружающей среде, в которой соответствующий организм существует (или мог бы существовать, в случае «концептуальных» геномов, полученных компьютерными методами). Однако, как только появились первые два полных бактериальных генома, вторым из которых был геном Mycoplasma genitalium [35], лишенной клеточной стенки паразитической бактерии с размером генома всего около 570 генов, возникла очевидная идея, что «истинный» наименьший набор можно естественным образом вывести, сравнивая геномы этих двух существенно различно специализированных бактериальных патогенов (Mushegian and Koonin, 1996b). Точнее, можно было бы ожидать, что ортологичные гены в двух организмах будет представлять собой набор основных биологических функций, которые необходимы для выживания клетки, независимо от уникального образа жизни каждого организма.
Сравнение геномных наборов H. influenzae и M. genitalium дало 240 пар ортологичных генов, охватывающих большую часть очевидно существенных клеточных функций. Тем не менее в этом консервативном наборе несколько важных функций явно отсутствовали. До сих пор мы не говорили о «настоящей биологии», о биологических функциях, ролях генов, но теперь мы должны начать думать биологически. Определение минимального набора основных биологических функций – задача непростая. Соблазнительно, конечно, попытаться «разобрать эволюцию по винтику»: идя от сравнительной геномики, определить минимальный набор основных генов, сохраняемых во всех клеточных формах жизни. Но этот подход упускает возможность, что разные организмы могли прийти к решению одной и той же принципиальной задачи независимыми путями. Мы увидим далее в этой главе, что такая гипотетическая возможность действительно отражает важный аспект биологической реальности. Таким образом, чтобы очертить минимальный набор клеточных функций, нам необходимо обратиться к логике биохимии и клеточной биологии. Знаний в этих областях несомненно достаточно, чтобы составить разумный каталог основных функций. Само собой, это знание несовершенно, поэтому на самом деле вычисление минимального набора генов требует многократного поочередного обращения к биологическому обоснованию и сравнительному геномному анализу. Мы с Аркадием Мушегяном предположили, что принципиально важные функции, отсутствующие среди 240 ортологов H. influenzae и M. genitalium, вероятно, исполняются неродственными или отдаленно родственными белками в этих двух бактериях. Мы привлекли определенные догадки, чтобы увеличить предполагаемый минимальный набор на 16 дополнительных генов M. genitalium (см. рис. 3-10). Этот простое упражнение в получении минимального набора генов соединением сравнительной геномики и биологической логики оказалось достаточно успешным и, по-видимому, определило приближенный функциональный репертуар простейшей бактериальной клетки, способной к самостоятельному росту в наиболее благоприятных условиях. В самом деле, последующие эксперименты с нокаутом генов подтвердили, что большинство из генов, включенных в минимальный набор, необходимы для выживания бактерий и что гены из минимального набора присутствуют в большинстве (хотя и не обязательно во всех) вновь секвенированных бактериальных геномах (Delaye and Moya, 2010; Koonin, 2003).
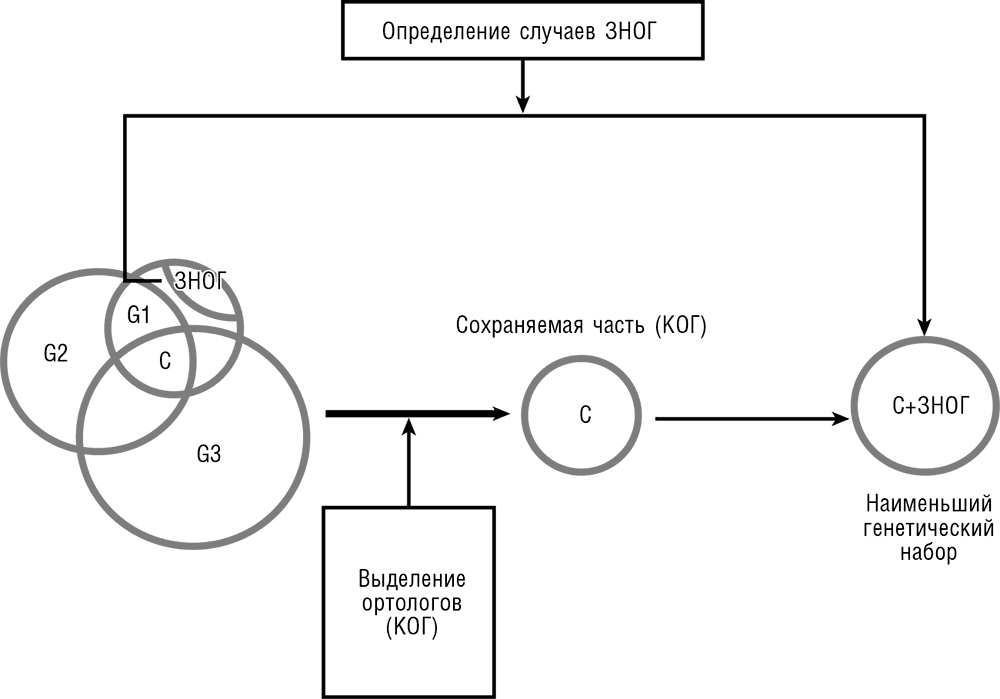
Рис. 3-10. Выделение минимального набора генов клеточной жизни методами сравнительной геномики. G1, G2, G3 – три сравниваемых генома; С – набор консервативных генов.
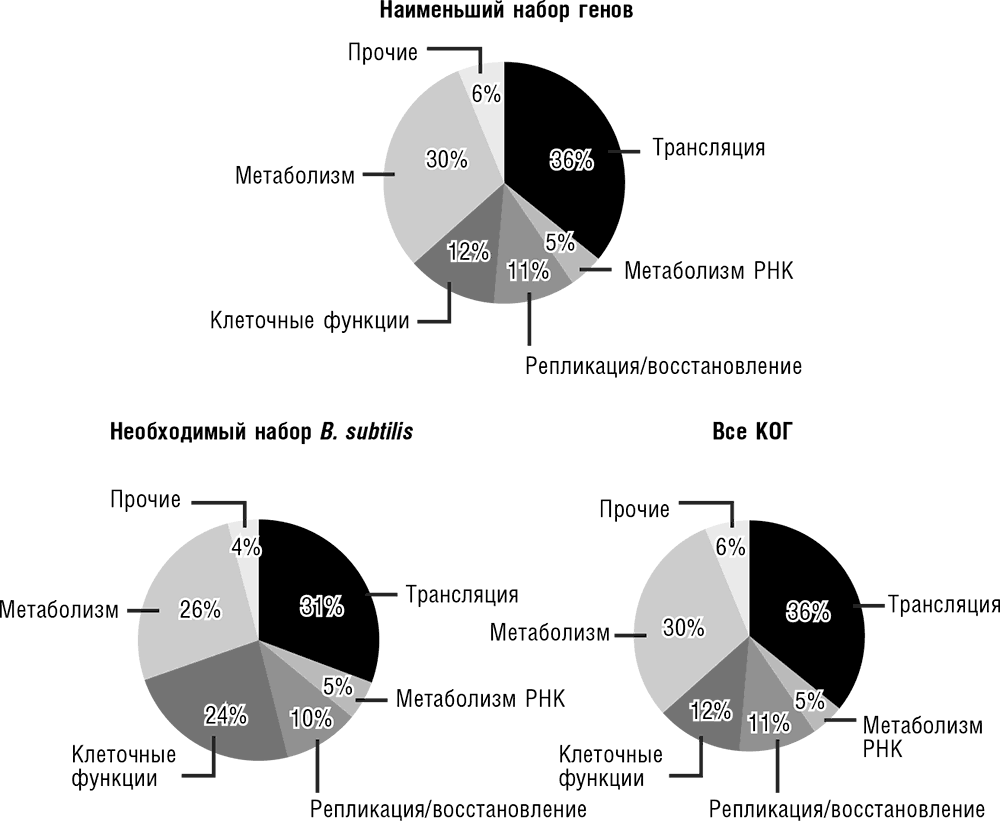
Рис. 3-11. Распределение числа генов по биологическим функциям в минимальном генном наборе, полном наборе КОГ и среди экспериментально определенных незаменимых генов бактерии Bacillus subtilis. Данные по Koonin, 2003.
Поучительно провести теперь функциональную перепись минимального бактериального набора генов. В этом наборе преобладают гены, которые кодируют белки, участвующие в передаче информации в клетке (репликации, транскрипции и, прежде всего, трансляции). Метаболические ферменты и белки транспортной системы представлены куда более разреженно, что вполне ожидаемо для организма, растущего в самой богатой из возможных сред. Этим минимальный набор генов резко отличается от полного набора КОГ, но напоминает набор незаменимых бактериальных генов (нокаут которых убивает бактерию, см. рис. 3-11). Эта особая эволюционная устойчивость систем передачи клеточной информации является одним из центральных обобщений сравнительной геномики. Мы вернемся к этому вопросу позднее.

Рис. 3-12. Эволюционный сценарий замещения неортологичных генов.
По-видимому, самым весомым результатом поиска минимального набора генов было открытие, что в списке легко обнаружимых ортологов недостает нескольких важных функций. Этот результат был подтвержден и значительно усилен при сравнении бактериальных геномов с первым архейным (Methanocaldococcus jannaschii), когда в наборе консервативных необходимых функций выявилось несколько дополнительных зияющих пробелов. Эти выводы были обобщены в понятии замещения неортологичных генов (ЗНОГ), эволюционном сценарии, при котором неродственные или отдаленно родственные гены (иными словами, не ортологи) становятся ответственными за одни и те же необходимые функции в разных организмах (Koonin, 2003). Сам этот эволюционный сценарий легко представить (см. рис 3-12): чтобы произошло ЗНОГ, эволюционирующая линия приобретает альтернативный, функционально избыточный ген для некоторой незаменимой функции и таким образом проходит через промежуточное состояние, в котором присутствуют обе реализации данной функции (такая избыточность часто наблюдается у организмов с более сложными геномами), а затем теряет начальный ген (Koonin and Mushegian, 1996). С ростом коллекции секвенированных геномов обнаруживается все больше организмов, в которых оба варианта действительно представлены для разнообразных функций; таким образом, сценарий эволюции ЗНОГ, представленный на рис. 3-12, становится все более правдоподобным.
В табл. 3–2 описывается несколько примеров ключевых биологических функций, для которых два или несколько неродственных ферментов по-разному представлены в частично дополнительных, но обычно перекрывающихся группах эволюционных линий. Даже эти отдельные примеры показывают, что ЗНОГ происходит в самых различных функциональных системах и путях. В дальнейшем, с заметным увеличением числа секвенированных геномов, стало ясно, что ЗНОГ и утрата генов в отдельных линиях настолько широко распространены, что лишь малое число функций являются действительно мономорфными и вездесущими (то есть представлены ортологичными генами во всех организмах). Вместе с тем универсальное ядро жизни уменьшилось почти до исчезновения: все, что остается универсальным, – это около тридцати генов белков трансляции и три больших субъединицы РНК-полимеразы, а также примерно равное число генов структурных РНК (рРНК и тРНК).
Таблица 3–2. Примеры замещения неортологичных генов.

Даже при исключении паразитических бактерий перечень универсальных генов расширяется незначительно (Koonin, 2003). Таким образом, за исключением небольшого числа генов, участвующих в основных этапах передачи информации, не существует универсального генетического ядра жизни, в связи с повсеместными ЗНОГ и потерей генов. Концепция небольшого, универсального набора функций, необходимых для поддержания клетки, остается жизнеспособной, но, учитывая комбинаторику ЗНОГ, этот наименьший набор функциональных ниш может заполняться огромным разнообразием генных ансамблей.
Date: 2016-05-15; view: 564; Нарушение авторских прав