Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Геномные ландшафты: распределение эволюционных ограничений по разным классам сайтов в геноме
|
|

Рис. 3–7. Схематические геномные ландшафты. Распределение эволюционных ограничений по разным сайтам в геномах прокариот и эукариот вскрывает различные принципы геномной архитектуры: а – геном прокариот; б – геном эукариот.
Любой геном может быть представлен в виде геномного ландшафта, графика, напоминающего панораму города, где каждому нуклеотидному сайту присваивается высота, пропорциональная силе влияющих на него эволюционных ограничений. Ограничения имеет смысл рассматривать как меняющиеся в диапазоне от 0 (лишенная ограничений, нейтрально эволюционирущая, функционально не значимая позиция) до 1 (полностью ограниченная, функционально важная позиция, в которой изменения недопустимы, см. рис. 3–7; Koonin and Wolf, 2010b). Распределения ограничений по геному значительно отличаются у форм жизни с различными архитектурами генома. Эти отличия проявляются особенно ярко, если говорить о сравнении, с одной стороны, вирусов и прокариот с их геномами «стена к стене», в основном состоящими из генов, кодирующих белок или РНК, и, с другой стороны, многоклеточных эукариот, в геномах которых кодирующие нуклеотиды находятся в меньшинстве (см. рис. 3–7). В пересчете на один сайт, ограничения в компактных геномах, особенно у прокариот, на несколько порядков сильнее, чем ограничения в больших геномах многоклеточных эукариот. Белок-кодирующие последовательности и последовательности, кодирующие структурные РНК, подвержены наиболее сильным ограничениям во всех геномах. Подавляющее большинство белок-кодирующих генов, особенно у прокариот, имеют низкие значения Ka / Ks, что указывает на сильное давление очищающего отбора на эти последовательности (см. рис. 3–8 и предыдущую главу). В то же время во всех группах организмов существует значительная положительная корреляция между Ka и Ks, указывающая, что даже синонимические сайты в белок-кодирующих генах ограничены примерно в пропорции к ограничениям на несинонимичные сайты (Drummond and Wilke, 2008; см. также гл. 4). Учитывая, что прокариотические геномы почти полностью состоят из белоккодирующих генов со вкраплениями генов структурных РНК и коротких межгенных промежутков, в основном занятых разно образно ограниченными регуляторными регионами, эти компактные геномы содержат мало неограниченных сайтов. Заметным исключением являются псевдогены, редкие у большинства прокариот, но распространенные у некоторых паразитических бактерий, особенно растущих внутри эукариотических клеток, например Rickettsia или Mycobacterium leprae (Harrison and Gerstein, 2002). Геномы большинства вирусов еще более компактны, чем геномы прокариот, причем почти вся последовательность генома занята белок-коди рующими генами.
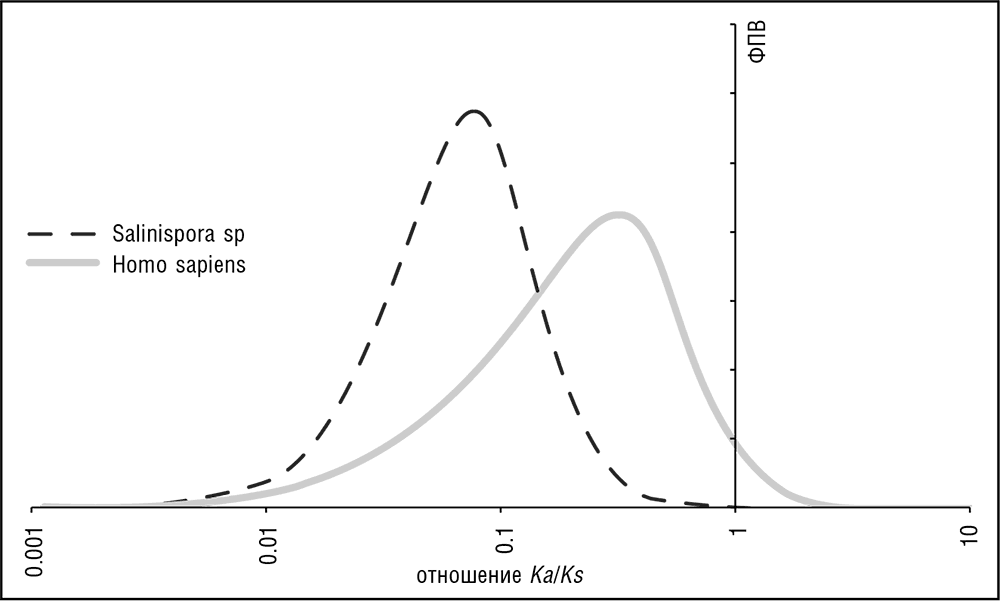
Рис. 3–8. Распределение отношения Ka / Ks в геномах прокариот и эукариот. Salinispora sp.: вычислено по ортологам в S. arenicola CNS-205 и S. tropica CNB-440 (актинобактерии). Homo sapiens: вычислено по ортологам в H. sapiens и Macaca mulatta (приматы). Значения Ка и Ks оценены с использованием программного обеспечения PAML (Yang, 2007). График в логарифмических координатах по оси абсцисс; ФПВ обозначает функцию плотности вероятности.
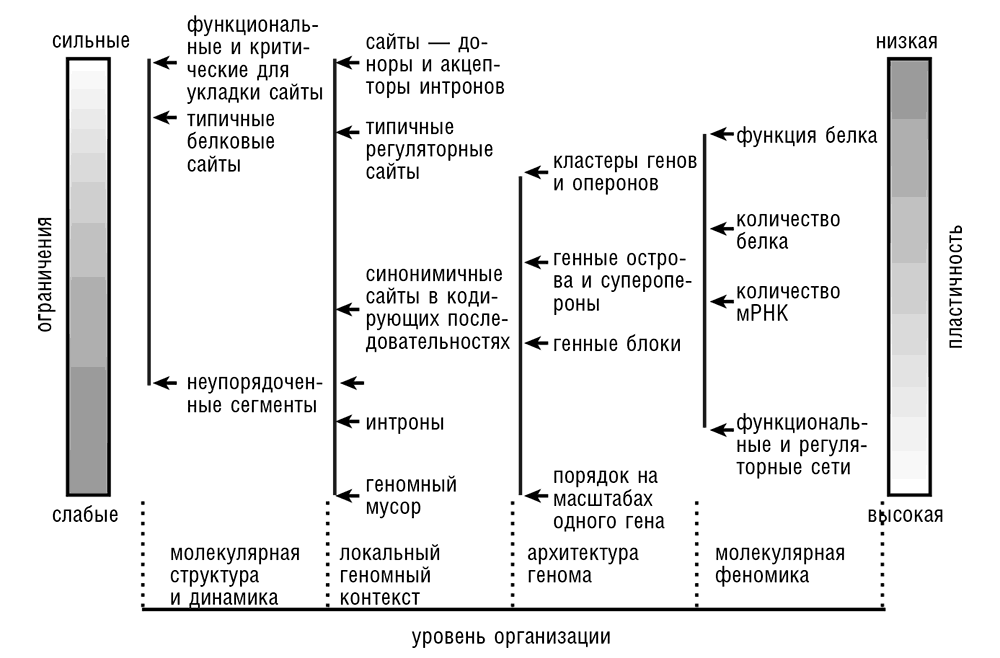
Рис. 3–9. Схематическая сводка эволюционных ограничений, действующих на различные классы геномных сайтов.
Одноклеточные эукариоты, напоминающие прокариот общей архитектурой генома, демонстрируют примерно одинаковые распределения эволюционных ограничений, хотя доля очевидно не подверженных ограничениям некодирующих последовательностей в их геномах несколько выше. Геномы многоклеточных эукариот (растений и особенно животных) являют собою разительную противоположность. Эти организмы имеют богатые интронами геномы с длинными межгенными промежутками; существенная, хотя и переменная часть этих некодирующих последовательностей, по-видимому, эволюционирует, не подвергаясь ограничениям. Доля нуклеотидов в геноме, подверженных эволюционным ограничениям, оценивается методами, основанными на критерии Макдональда – Крайтмана (см. табл. 2–2). Полученные оценки существенно отличаются даже между животными: у Drosophila около 70 процентов нуклеотидных сайтов в геноме, в том числе 65 процентов некодирующих участков, по всей видимости, подвержены отбору (в том числе положительному), а у млекопитающих эта доля оказывается в интервале 3–6 процентов (Koonin and Wolf, 2010b). Примечательно, однако, что абсолютное число подверженных отбору сайтов в столь разных по размеру геномах этих животных довольно близко. Напротив, в Arabidopsis, растении с геномом, размером и общей архитектурой сравнимыми с таковыми Drosophila, доля некодирующих подверженных ограничениям участков, по-видимому, существенно ниже.
Резюмируя существующее понимание ограничений, влияющих на различные классы и сайты во всем известном разнообразии геномов (см. рис. 3–9), отметим, что некоторые фундаментальные простые выводы являются бесспорными. В частности, нет никаких сомнений, что несинонимичные сайты в белок-кодирующих последовательностях и последовательности, кодирующие структурные РНК, являются одними из наиболее сильно ограниченных во всех геномах и что характерное распределение ограничений (геномный ландшафт) сильно коррелирует с архитектурой генома (Koonin and Wolf, 2010b). Однако помимо этих основных принципов, и довольно неожиданно, оказывается, что эволюционные режимы сильно различаются даже для некоторых относительно близких таксонов, таких как членистоногие и позвоночные. Чтобы выработать всеобъемлющую картину эволюционных ограничений и давления, формирующих геном, требуется еще множество дополнительных исследований по различным организмам. В последующих главах рассматриваются различные проявления давления отбора, влияющие на разные части генома.
Вселенная генов
Интеграция результатов сравнительной геномики позволяет нам начать строить карту всей «вселенной генов». Глобальная эволюционная устойчивость генов, проявляющаяся прежде всего в сохранении белковых и РНК-последовательностей, стала очевидной в результате самых первых сравнений секвенированных прокариотических и эукариотических геномов: бактерии Haemophilus influenzae и Mycoplasma genitalium, археи Methanocaldococcus jannaschii и эукариотических дрожжей Saccharomyces cerevisiae (Tatusov et al., 1997). Ключевое обобщение сравнительной геномики состоит в том, что гены не просто сохраняются на различных интервалах эволюционного процесса, но и представляют собой дискретные единицы эволюции, а именно ортологичные эволюционные линии (см. табл. 3–1). В сегодняшней коллекции секвенированных геномов найдены ортологи в далеких таксонах для значительного большинства белок-кодирующих генов в каждом геноме. Ярким примером являются недавние результаты секвенирования генома примитивных животных: многочисленные гены Trichoplax и губки связаны ортологичными отношениями с генами млекопитающих и птиц (Putnam et al., 2007; Srivastava et al., 2008; Srivastava et al., 2010). Один из выводов состоит в том, что характерная продолжительность жизни животного гена в этих линиях охватывает по меньшей мере сотни миллионов лет. Многие другие группы животных, такие как насекомые, утеряли многочисленные гены (Koonin et al., 2004), так что судьба одного и того же гена в большинстве случаев отличается в разных линиях, в результате чего мы получаем «пятнистую» филетическую модель. (Как подчеркивается далее в этой главе, множество поистине универсальных генов чрезвычайно мало.) Судьбы конкретных генов в разных линиях зависят как от случайных факторов, так и от различий в давлении отбора (см. гл. 9). Результаты обширного сравнительного анализа геномов растений, грибов и прокариот полностью совместимы с этим выводом. Когда гены в геноме классифицируются по их относительному «возрасту» (то есть филогенетической глубине, на которой обнаруживаются гомологи), наблюдаемое расхождение подобно для удаленных друг от друга организмов, как показано на рис. 3–3 для генетических наборов человека и грибка Aspergillus fumigatus (Wolf et al., 2009), двух организмов, разделенных, по-видимому, миллиардами лет эволюции. Тем не менее распределения генных возрастов поразительно похожи: в каждом случае древних генов, для которых легко обнаруживаются гомологи в далеких таксонах, значительно больше, чем «молодых» генов. Несмотря на частую потерю в отдельных эволюционных линиях, гены характеризуются чрезвычайной долговечностью, и многие из них, возможно, бессмертны [34].
Как обсуждается далее в этой книге (гл. 5 и 7), пути передачи генетической информации у прокариот принципиально отличаются от таковых у эукариот. Тем не менее доли консервативных генов у них примерно равны. В настоящее время эта доля хорошо известна и очень близка у разнообразных бактерий и архей, почти как фундаментальная постоянная: для 70–80 процентов генов ортологи обнаруживаются в далеких организмах (Koonin and Wolf, 2008b; см. рис. 3–4).
Date: 2016-05-15; view: 699; Нарушение авторских прав