Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Древние гены составляют в геноме большинство и имеют отчетливую эволюционную судьбу
|
|
Как уже упоминалось в предыдущей главе, Эрнст Майр, великий эволюционист XX века и один из основателей СТЭ, с уверенностью предсказывал исходя из больших фенотипических различий между организмами, что гены разных организмов, даже близкородственных, не будут иметь узнаваемого сходства. Ошибочность этого предсказания оказалась просто феерической, что само по себе делает его нетривиальным и ценным. Сравнение последовательностей даже в догеномный период выявило высокий консерватизм последовательностей у некоторых гомологичных белков и молекул некодирующих РНК по всему спектру жизни, от бактерий до млекопитающих (см. предыдущую главу). Более того, высокая степень сходства последовательностей существует у древних паралогов, которые, по-видимому, происходят от копий, ведущих свое происхождение от LUCA (Gogarten et al., 1989; Iwabe et al., 1989). Геномика позволяет перевести это общее понимание в количественное разбиение генов любого генома на классы эволюционной консервативности (см. рис. 3–3; Koonin and Wolf, 2008b).
Ключевое открытие сравнительной геномики состоит в том, что большинство генов в каждом геноме могут считаться высококонсервативными – они имеют легко обнаруживаемые гомологи в организмах, разделяемых сотнями миллионов лет эволюции (например, в случае генов человека, на уровне общего предка позвоночных; см. рис. 3–3; Wolf et al., 2009). Это открытие демонстрирует поразительную устойчивость последовательностей РНК и белков в процессе эволюции: типичное время исчезновения сходства последовательностей у гомологичных генов сравнимо со временем существования жизни на Земле. Помимо основополагающего значения, данный факт имеет огромные практические последствия: благодаря ему, прежде всего, сравнительная геномика становится крайне информативной и действенной.
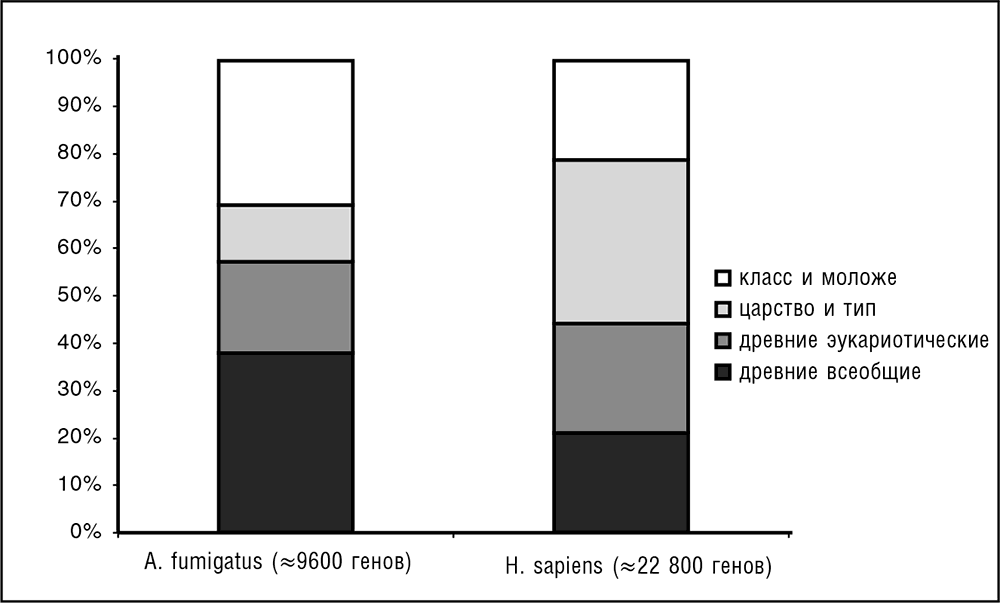
Рис. 3–3. Распределение генов по эволюционному возрасту. «Эволюционный возраст» соответствует самому старому таксономическому узлу, в котором могут быть определены гомологи для белка, производимого данным геном. В частности, для человека древние всеобщие означает «гомологи, обнаруживаемые у прокариот», древние эукариотические означает «гомологи, обнаруживаемые у прокариот вне супергруппы униконтов» (см. гл. 7), царство и тип означает «гомологи, обнаруживаемые у животных вне класса млекопитающих», а класс и моложе означает «вне класса млекопитающих гомологи не обнаружены» (данные по Wolf et al., 2009)
Структуру эволюционного процесса определяют не только консервативные последовательности. На протяжении чрезвычайно длительных эволюционных периодов не просто сохраняется сходство последовательностей РНК и белков, но и гены имеют свойство сохранять свою уникальность. Иными словами, большинство генов развиваются как ортологичные линии, с редкими случаями дупликации (Koonin, 2005). Устойчивость ортологии генов становится очевидной благодаря простой процедуре, широко применяемой в сравнительной геномике и позволяющей эффективно выявлять ортологичные наборы генов. При этом ортологи обнаруживаются как «наилучшие совпадения при двунаправленном сравнении» (bidirectional best hits): все закодированные в геноме белковые последовательности сравниваются со всеми белками, закодированными в другом геноме, a затем процедура повторяется в обратном направлении (Tatusov et al., 1997). Пары генов, дающие наилучшие совпадения (те, которые демонстрируют наибольшее сходство последовательностей) при обоих направлениях сравнения, считаются возможными ортологами; нетрудно применить эту процедуру к нескольким видам путем совмещения треугольников двунаправленных совпадений, имеющих общую сторону (см. табл. 3–1). Примечательно, что такой прямолинейный подход в большинстве случаев хорошо срабатывает: к примеру, порядка 70 процентов генов организмов, разделенных приблизительно 100 миллионами лет эволюции, таких как люди и мыши, легко идентифицируются как ортологи при помощи описанной процедуры (Wolf et al., 2009). Если применить простую модификацию этого алгоритма и включить дупликации генов, характерных для одной линии наследования (дупликации, образовавшиеся после расхождения сравниваемых видов), такой подход позволяет идентифицировать наборы ортологов (известных как кластеры ортологичных генов, КОГ) во многих геномах, в том числе столь удаленных друг от друга, как археи и бактерии – представители двух доменов прокариот (см. гл. 5). Более точные и мощные способы обнаружения ортологов требуют подробного анализа филогенетических деревьев (см. табл. 3–1); впрочем, результаты такого анализа обычно близки к тем, что дают более простые методы, основанные только на сравнении последовательностей. Разумеется, для части генов история дупликаций и потерь настолько сложна, что обнаружить КОГ трудно, поэтому они становятся нечеткими кластерами с неопределенной внутренней структурой. По счастью, этих «трудных» генов в каждом геноме относительно немного.
Таблица 3–1. Классификация гомологичных связей генов: ортологи, паралоги и методы их определения.
Эволюционные связи генов:
• Гомология: гены, имеющие общее происхождение.
• Ортология: гомологичные гены, эволюционировавшие путем видообразования.
• Паралогия: гомологичные гены, эволюционировавшие путем дупликации.
• Ксенология: гомологичные гены, имитирующие ортологи, но образовавшиеся в результате горизонтального переноса гена из другой ветви.
• Паралогия, внутренняя и внешняя: паралогичные гены, возникшие в результате видоспецифической дупликации после (внутренняя) или до (внешняя) определенного события видообразования.
• Со-ортология: внутренне-паралогичные гены, совокупно ортологичные по отношению к генам другой ветви (из-за их общего происхождения в ходе видообразования).
• Ортологичная группа (КОГ): совокупность всех потомков данного предкового гена.
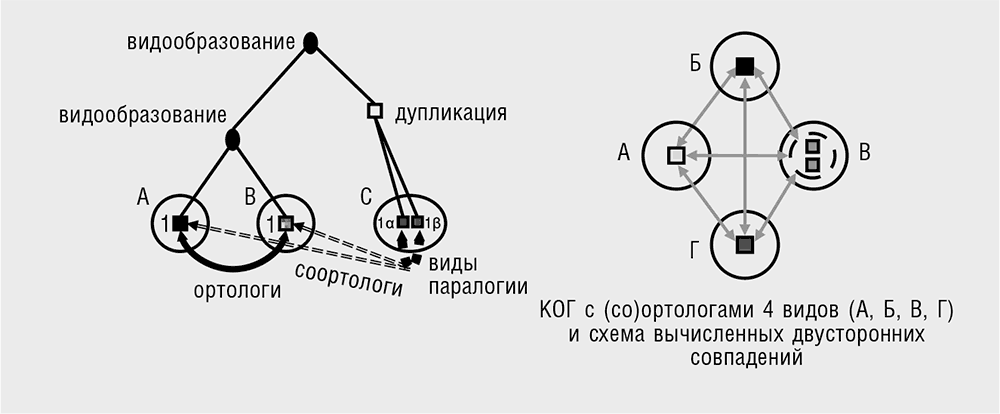
Изначально не вполне складная аббревиатура КОГ относилась к кластерам ортологичных групп (белков), чтобы обозначать соортологичные связи, вызванные дупликацией генов (см. табл. 3–1; Tatusov et al., 1997). Сейчас я предпочитаю расшифровывать КОГ просто как кластеры ортологичных генов, однако само по себе это сокращение остается чрезвычайно удобным для обозначения фундаментального свойства таких кластеров. Эта трехбуквенная аббревиатура широко используется в литературе, и я использую ее в данной книге в качестве сокращенного названия наборов ортологичных генов. Обычно каждый секвенированный геном более чем на 70 процентов состоит из генов, относящихся к КОГ (см. рис. 3–4). В эволюции генома, к которой мы обращаемся в этой книге неоднократно, эта величина представляется важной. Таким образом, существенное большинство генов в каждом геноме весьма консервативно, то есть представлено ортологами во многих далеко отстоящих друг от друга организмах.
Date: 2016-05-15; view: 598; Нарушение авторских прав