Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Диагностика и типирование BLV провируса, выявленного у крупного рогатого скота краснодарского края
|
|
Новосибирский государственный аграрный университет
Ключевые слова: вирус лейкемии крупного рогатого скота, генотипирование, ПДРФ-анализ, LTR-область, болезни животных, вирусы, биотехнология животных.
Аннотация: На протяжении многих лет изучение BLV является актуальной проблемой. Так как до настоящего времени не выявлены многие биологические особенности течения вируса. Мы проанализировали последовательности LTR-области 11 изолятов BLV, в том числе циркулирующих на поголовье скота на территории Российской Федерации. Подобрали наиболее специфичные и чувствительные праймеры, фланкирующие наиболее вариабельный участок LTR-области.
Key words: bovine leukemia virus, genotyping, RFLP analysis, LTR-region, animal diseases, viruses, animal biotechnology.
Abstract: Formanyyearsthe study of BLV is an actual problem. So far not revealed many of the biological characteristics of the flow of the virus. We analyzed sequences of LTR-region 11 of BLV isolates, including circulating in livestock population on the territory of the Russian Federation. Picked up the most specific and sensitive primers flanking the most variable plot LTR-region.
Итак, проводя сравнительные исследования крупного рогатого скота с разной степенью компрометации к лейкозу (инфицированные BLV в стадии бессимптомного вирусоносительства; в развернутой гематологической стадии; и интактные к BLV животные), а также при исследовании иммунодепрессивного эффекта BLV на организм вирусоносителя, нам представлялось актуальным выявить степень генотипической однородности BLV провируса [1-6]. Это можно было достичь, используя методику, в частности RealTimeSYBRgreen для изучения структурного разнообразия провируса по нуклеотидному составу на основании температурных пиков.
Необходимо отметить, что в любом организме хозяина хронически персистирующий вирус может претерпевать мутационные изменения, направленные на увеличение патогенности, на преодоление более эффективных защитных механизмов хозяина [1, 3, 4]. Поэтому, наряду с генетическим статусом макроорганизма, во внимание необходимо принимать и генетическую изменчивость самого патогена, который, возможно, в процессе длительной персистенции способен изменять свою патогенность, в сторону, как комменсализма, так и усилении япатогенных свойств [5-7]. Генетическая вариабельность BLV у инфицированного крупного рогатого скота достаточн охорошо охарактеризована.
На основании иметода ПДРФ и ДНК секвенирования вируса лейкоза крупного рогатого скота классифицировано три подгруппы вируса - бельгийская, австралийская и японская [1, 4, 6]. Так как секвинирование достаточно затратный метод генотипирования, для распределения выборки по известным генотипам применяют ПДРФ-анализ (полиморфизм длин ристрикционных фрагментов) или методику RealtimePCR с флуоресцентно-мечеными олигонуклеотидными зондами. Наиболее широко используется 5'-экзонуклеазный метод, основанный на эффекте пространственного разделения флуорофора и гасителя флуоресценции вследствие расщепления Taq ДНК-полимеразой зонда, связанного с амплифицируемой последовательностью ДНК [4].
Целью наших исследований явилось проведение генотипирования BLV-провируса по env гену циркулирующего на поголовье крупного рогатого скота Краснодарского края, с распределением исследуемых образцов по структурной составляющей на группы по gag гену. В связи с этим были поставлены следующие задачи: 1) выделить ДНК из крови серопозитивных животных; 2) провести ПЦР в реальном времени; построить графики температур плавления и накопления продуктов амплификации; 3) провести биоинформационный анализ полученных данных.
Объектом исследований являлся крупный рогатый скот, принадлежащий ПЗ «им. Чапаева» Краснодарского края, на которых мы и раннее проводили исследования. Материалом исследования послужила кровь РИД - положительных животных. Выделение ДНК из крови производили сорбентным методом с использованием наборов МедиГен (Новосибирск).
Для определения вирусоносителя по гену env gp51 применили следующие праймеры: № 1. 5´TCTGTGCCAAGTCTCCCAGATA-3´, № 2. 5´СССACAAGGGCGGCGCCGGTTT-3´ [6-9], для определения вирусоносителя по гену gag p24 применили следующие праймеры (347 bp): № 1. 5’GGAGGWGGRAAGATGCGAACTATT 3’, № 2. 5’GTCCGYTCTA CYAACCCTGAACTT 3’ [4, 6], синтезированные ООО «Медиген». Применяли способ проведения PCR realtime с SYBRgreen-II.
На рисунке 1 изображен график ПЦР real-time, в котором отображается накопление специфических продуктов амплификации BLV (CRE-область).

Риcунок 1 – Постановка ПЦР реакции с праймерами CRE-область
После адаптации режимов амплификации, на рисунке 2 мы видим более четкое отображение кривых накопления продуктов амплификации.

Риcунок 2 – Постановка ПЦР реакции с праймерами CRE-область, после адаптации температурно временных параметров
Выход кривых плавления примерно в одинаковых температурных пределах. Подбор оптимальной температуры плавления осуществляли эмпирическим путем, в трех разведениях ДНК. На рисунке 3 хорошо визуализируются как положительные образцы, так и отрицательный контроль.

Риcунок 3 – Кривая плавления продуктов амплификации, (амплификоны CRE-область)
Как мы можем заметить максимальное количество образцов было отмечено при температуре от 76 до 77º С. Однако, температурная кривая плавления некоторых проб отличалась.
На следующем графике (рис. 4) изображены кривые температур плавления амликонов (env).
На графике 5 изображен участок с наибольшей изменчивостью (t 75-95 ºС). Следует отметить, что для данного аплифицируемого участка выборки оптимальная температура плавления составляет от 79,6 до 81,4ºС. Оптимальные температуры плавления и накопления продуктов амплификации определяли опытным путем, режимы аплификации адаптировали.
 Рисунок 4 – Кривая плавления продуктов амплификации, ДНК gag гена (провируса BLV) ° С
Рисунок 4 – Кривая плавления продуктов амплификации, ДНК gag гена (провируса BLV) ° С
|  Рисунок 5 – Кривая плавления продуктов амплификации участка ДНК gag гена (провируса BLV), ° С
Рисунок 5 – Кривая плавления продуктов амплификации участка ДНК gag гена (провируса BLV), ° С
|
 Рисунок 6 – Кривая подборки температур плавления продуктов амплификации, участка ДНК gag гена (провируса BLV) ° С
Рисунок 6 – Кривая подборки температур плавления продуктов амплификации, участка ДНК gag гена (провируса BLV) ° С
|  Рисунок 7 – Участки распределения исследуемых образцов на группы по структуре ДНК (участок 444 п.н. env ген)
Рисунок 7 – Участки распределения исследуемых образцов на группы по структуре ДНК (участок 444 п.н. env ген)
|
Далее был проведен более точный анализ - НРМ (по кривым плавления), результаты которого показали нам, что генотипы опытных образцов напрямую зависят от нуклеотидного состава. Конкретный профиль плавления, который отражает кривые плавления, определяется последовательностью оснований в исследуемой ДНК. При нагревании до определенной температуры связи между нитями ДНК разрываются, и высвобождается краситель, на который реагирует прибор. Пики на дифференциальных кривых плавления связаны с «выплавлением» участков молекулы в интервале в несколько десятых градуса отдельных с характерным размером в несколько сотен пар оснований.
Различия изменения температур плавления не весомы а, соответственно, данные образцы можно сделать вывод об относительной однородности структуры исследуемых образцов, в то время как наблюдались значительные структурные различия на другом исследуемом участке ДНК провируса (gag) этих же образцов (Рис. 6-8). На графике хорошо читаются температурные пики, по которым группу распределили на несколько групп по нуклеотидному составу. Так, данные образцы были распределены на пять групп, с температурами плавления от 64 до 88 °С:
1 - температура плавления 64-66°С;
2 - 66,1-68°С;
3 - 82-84°С;
4 - 84,1-86°С;
5 - 86,1-88°С.
Самой многочисленной оказалась 5 группа, с существенным преобладанием, далее по убыванию третья, вторая, первая и четвертая. Проведя биоинформационный анализ данных образцов, мы определили, что данные группы соответствуют международной классификации генотипов (I-VII).
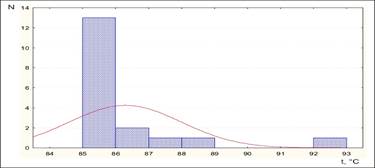
Рисунок 8 – Распределение исследуемых образцов на группы по структуре ДНК (участок 347 п.н. gag ген)
Анализируя полученные данные по температурным кривым образцов исследуемых по gag гену BLV (провируса), следует отметить, что образцы также распределились на 5 групп, однако совершенно в других температурных рамках.
Так, данные образцы были распределены также на пять групп, однако при других рамках температур плавления от 85 до 93 °С:
1 - 85-86 °С;
2 - 86, 1-87 °С;
3 - 87,1-88 °С;
4 - 88,1-89 °С;
5 - 92-93 °С.
Характеризуя данные группы, напрашивается вывод об определенной гомогенности 1-4 групп, и совершенно отличающейся температурами плавления образцов, входящих в 5 группу. Данный факт указывает на большое содержание в структуре ДНК, С (цитозин) и G (гуанин), которые способны образовывать тройные связи, а, соответственно, при их плавлении необходимы более высокие температуры. Так как на данный момент не существует определенной классификации генотипов, по данному участку gag гена, в дальнейшем предстоит провести секвенирование нескольких образцов из каждой группы.
Биоинформационным анализом результатов исследований ДНК провирусов BLV от крупного рогатого скота Краснодарского края выявлено 5 групп паттернов.
Распределение выборки, по температуре плавления, неравномерное. Наиболее многочисленной оказалась группа – 86, 1-880; далее в хронологическом порядке – 82 -840; 66, 1-68; 64 – 880 и 84,1 – 860 по gag гену.
Список литературы:
1. GilletN, FlorinsA, BoxusM, BurteauC, NigroA, etal. (2008) Mechanism of leukomogenesis induced by bovine leukemia virus: prospect for novel anti-retroviral therapies in human. Retrovirology 4: 18. doi: 10.1186/1742-4690-4-18
2. Kamoi, K., Yamamoto, K., Misawa, A., Miyake, A., Ishida, T., Tanaka, Y., Mochizuki, M., and Watanabe, T. (2006). SUV39H1 interacts with HTLV-1 Tax and abrogates Tax transactivation of HTLV-1 LTR. Retrovirology 13, 3-5.
3. Mansky LM, Temin HM (1994) Lower mutation rate of bovine leukemia virus relative to that of spleen necrosis virus. J Virol 68: 494–499.
4. Meas S., Usui T., Ohashi K et al. Vertical transmission of bovine leukemia virus and bovine immunodeficiency virus in dairy cattle herds / Meas S., Usui T., Ohashi K., Sugimoto C., Onuma M. // Veterinary Microbiology. 2002. Т. 84. № 3.С. 275-282.
5. Gillet N., Sanchez-Alcaraz T., Willems L. // Philosophical Transactions of the Royal Society of London. SeriesB: BiologicalSciences. 2006. Т.
6. Батенёва Н.В., Смирнов П.Н., Михнович И.В. Изучение распространения условных генотипов вируса лейкоза крупного рогатого скота // Сельскохозяйственная биология. 2012. № 4. С. 69-72
7. Дробот Е.В. Результаты изучения генотипического разнообразия вируса лейкоза крупного рогатого скота и особенности эпизоотологического и гематологического проявления. Автореф. канд. дис. Новосибирск, 2007.
8. Петропавловский М.В. Региональная молекулярно-генетическая структура вируса лейкоза крупного рогатого скота / Петропавловский М.В., Донник И.М. //Ветеринария Кубани. 2010. № 3. С. 12-13.
9. Смирнов П.Н. Болезнь века -лейкоз крупного рогатого скота. -Новосибирск, 2007.
Date: 2015-12-10; view: 546; Нарушение авторских прав