Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Периферические
|
|
Поверхностные белки (пб). Эти белки располагаются на внешней и внутренней поверхностях мембраны, удерживаясь преимущественно электростатическими силами. Такие белки занимают 75-80 % поверхности. Белковые молекулы покрывают мембрану с обеих сторон и придают ей эластичность и устойчивость к механическим повреждениям.
Интегральные белки (иб). Эти белки могут пронизывать двойной слой липидов насквозь. Такие белки являются главным компонентом, ответственным за избирательную проницаемость клеточной мембраны. Некоторые из них (к) образуют систему селективных каналов (пор) или функционируют как ионные насосы и регулируют, например, электрохимическую систему возбуждения клетки. Диаметр каналов составляет 0,35-0,8 нм. Количество их относительно невелико (например, в эритроцитах вся площадь каналов составляет 0,06 % от площади поверхности). Полярные группы молекул белков в каналах направлены в сторону отверстия каналов, а неполярные вступают во взаимодействие с молекулами липидов. Стенки каналов обладают электрическими зарядами.
Существуют 4 способа ассоциации белков с липидным слоем:
· интегральные белки, пронизывающие мембрану насквозь
· поверхностные белки, погруженные в липидном бислое частично
· белки, удерживающиеся нековалентными взаимодействиями с другими мембранными белками
· белки, ковалентно соединенные с одной или двумя цепями жирных кислот. 
- Физические свойства и фазовые переходы в мембранах
Внутреннее давление=100 Па
Вязкость=30-100 млПа
σ 0,03-3 Н/м
С=0,5-1,3*10^(-6) Ф
Е1=2-2,2
Е2=10-20
R=1 см^2
Плотность липидного бислоя составляет 800 кг/м3, что меньше, чем у воды.
Размеры. По данным электронной микроскопии, толщина мембраны (L) варьирует от 4 до 13 нм, причем различным клеточным мембранам присуща разная толщина.
Прочность. Предел прочности на разрыв для мембраны низок. В условиях организма средние деформации составляют около 0,01 %. Чтобы довести мембрану до разрыва, достаточно внутреннего давления 100 Па. Живая клетка может осуществлять осморегуляцию только за счет изменения своей формы, но не за счет растяжения мембраны.
Деформируемость. Клеточная мембрана легко подвергается деформации сдвига. Например, в потоке эритроцитов с градиентом скорости происходит вращение мембраны вокруг содержимого клетки. Мембрана обладает высокой гибкостью. При оценке механических свойств мембраны эффективный модуль упругости принимается равным 0,45 Па.
Вязкость. Липидный слой мембраны имеет вязкость η = 30-100 мПас (что соответствует вязкости растительного масла).
Поверхностное натяжение равно 0,03-3 мНм-1, что на 2-3 порядка ниже, чем у воды (73 мНм-1).
Коэффициент проницаемости мембранного вещества для воды равен 25-33х10-4 см/с.
Мембрана - конденсатор. Двойной фосфолипидный слой уподобляет мембрану плоскому конденсатору, обкладки которого образованы электролитами внеклеточного и внутриклеточного (цитоплазмы) растворами с погруженными в них поверхностными белками и головками липидных молекул. Обкладки разделены диэлектрическим слоем, образованным неполярной частью липидных молекул - двойным слоем их хвостов. Электроемкость 1 см2 мембраны составляет 0,5-1,3 мкФ. Напряженность электрического поля в мембране составляет приблизительно 20х106 В/м.
Диэлектрическая проницаемость мембраны составляет: для фосфолипидной области ε = 2,0-2,2; для гидрофильной области ε = 10-20.
Электросопротивление 1 см2 поверхности мембраны составляет 102-105 Ом (что в десятки миллионов раз больше сопротивления внеклеточной жидкости или цитоплазмы). Электроизоляционные свойства мембраны значительно превосходят свойства технических изоляторов.
Жидкокристаллическое состояние. Молекулы в мембране размещены не беспорядочно, в их расположении наблюдается дальний порядок. Фосфолипидные молекулы находятся в двойном слое, а их гидрофобные хвосты приблизительно параллельны друг другу. Есть порядок и в ориентации полярных гидрофильных головок. Физическое состояние, при котором есть дальний порядок во взаимной ориентации и расположении молекул, но агрегатное состояние жидкое, называется жидкокристаллическим состоянием.
Строго говоря текучесть мембраны ограничена внутренней гидрофобной фазой, которая состоит из углеводородных цепей жирных кислот. Эта фаза, однако, не всегда бывает жидкой. При охлаждении до температур ниже 10оС мембраны замерзают, т.е. жидкая фаза затвердевает, приобретая свойства двумерного кристалла.
В мембранах, образованныых синтетическими липидами, фазовый переход из жидкого в твердое состояние может происходить при более высоких температурах, в зависимости от химического состава фосфолипида.
При нормальных физиологических условиях мембраны находятся в жидком состоянии, однако, в отличие от жидкостей, они имеют строгую упорядоченную структуру, поэтому это состояние мембран называется жидкокристаллическим. По данным физических методов биологические мембраны способны испытывать фазовый переход первого рода. С понижением температуры мембраны переходят из жидкокристаллического состояния (золь) в твердокристаллическое (гель). При таком переходе сохраняется общая структура мембраны, но порядок в системе еще более возрастает: «хвосты» липидов вытянуты строго параллельно друг другу, их колебания ограничены. Если в золь-состоянии площадь мембраны, приходящаяся на одну молекулу липида, составляет 0,58 нм2, то в гель-состоянии эта величина уменьшается до 0,48 нм2. Толщина мембраны при переходе в гель-состояние увеличивается, но за счет уменьшения площади объём мембраны в целом уменьшается.
Подвижность липидных молекул в обоих фазовых состояниях существенно отличается. В гель-состоянии липиды способны совершать только совместные колебания или вращательные движения. В жидком состоянии липидные «хвосты» имеют большую свободу, особенно велика их подвижность в середине мембраны.
Молекулы липидов жидкой мембраны могут находиться в в двух конформациях: в полностью транс- конформации и в гош-транс-гош-конформации. В гель-состоянии реализуется только транс-конформация. Длина молекулы в гош-конформации меньше, чем в транс-, а ширина, наоборот, больше. Этим объясняется уменьшение толщины и увеличение площади мембраны, приходящейся на один липид, при переходе из гель- в золь-состояние. Способность липидов жидкокристаллической мембраны переходить в гош-конформации способствует повышению её проницаемости для некоторых гидрофильных низкомолекулярных соединений. Если два соседних участка рядом расположенных молекул находятся в гош-конформации, то образуется так называемый кинк. Кинки способны перемещаться вдоль липидных «хвостов». Если небольшая молекула, окажется между двумя кинками, то она сможет пересечь гидрофобный слой.
В последнее время доказано, что при фазовых переходах из золь-состояния в гель- и обратно в мембранах образуются поры диаметром 2-6 нм. То есть при фазовом переходе увеличивается проницаемость мембраны для ионов и низкомолекулярных соединений. Так, проведение нервного импульса определяется изменением проницаемости мембраны для некоторых ионов. Существует предположение, что механизмы терморецепции, также определяются фазовым переходом в мембранах рецепторов. И ещё, так как фазовый переход может иметь и химическую природу (например, липиды мембраны могут перейти в гель-состояние вследствие увеличения концентрации ионов кальция в межклеточной жидкости), то, возможно, эти процессы определяют и хеморецепцию.
Фазовый переход в мембранах происходит не мгновенно, а на протяжении некоторого температурного интервала. Температурой фазового перехода называется та температура, при которой одна половина мембранных липидов находится в золь-состоянии, а другая половина в гель-состоянии. Температура фазового перехода зависит от липидного состава мембран: чем больше в «хвостах» липидов двойных связей (а их число в одном «хвосте» может колебаться от 0 до 4), то есть чем больше степень ненасыщенности жирнокислотных остатков, тем ниже температура фазового перехода. Для мембран, состоящих из насыщенных липидов, эта величина составляет +60  С, а для мембран, состоящих из ненасыщенных липидов, - снижается до -20 С. Такая разница в температурах плавления объясняется тем, что ненасыщенные жирные кислоты, находящиеся в цис-конформации, способствуют удалению друг от друга молекул липидов, и, следовательно, уменьшают силы сцепления между ними. Чем слабее связь между молекулами, тем ниже температура перехода мембран в жидкую фазу.
С, а для мембран, состоящих из ненасыщенных липидов, - снижается до -20 С. Такая разница в температурах плавления объясняется тем, что ненасыщенные жирные кислоты, находящиеся в цис-конформации, способствуют удалению друг от друга молекул липидов, и, следовательно, уменьшают силы сцепления между ними. Чем слабее связь между молекулами, тем ниже температура перехода мембран в жидкую фазу.
Живые организмы, обитающие в разных климатических условиях, имеют различное соотношение насыщенных и ненасыщенных связей в молекулах липидов, что обеспечивает им приспособление к холоду или жаре в зависимости от среды обитания. Более того, это соотношение меняется для разных участков тела одного и того же организма. Например, температура ноги возле копыта полярного оленя может составлять -20 С, а температура ноги возле туловища достигает +30 С. Однако клеточные мембраны не претерпевают фазовый переход за счет того, что мембраны клеток возле копыта содержат больше ненасыщенных липидов, а возле туловища – больше насыщенных. Известно, что животные жиры при комнатной температуре обычно находятся в твердом состоянии, а растительные, наоборот, - в жидком. Это объясняется тем, что в животных клетках меньше ненасыщенных липидов, чем в растительных.
Фазовые переходы мембранных липидов носят кооперативный характер, то есть чем большее количество липидов совершило фазовый переход, тем легче это будет сделать остальным. В идеале, если на каком-либо участке одна молекула липида перешла из твердой фазы в жидкокристаллическую, то за этим сразу же последует фазовый переход всех липидов этого участка (закон «всё или ничего»). Такой участок называется кооперативной единицей, а число молекул, входящих в него, - размером кооперативной единицы n. Для липидных мембран n может достигать нескольких десятков. Величина n тем больше, чем более однороден химический состав мембран. Для уменьшения однообразия липидного состава мембраны животных клеток содержат большое количество холестерина. Например, в эритроцитах млекопитающих его удельная доля от всех липидов составляет 40 - 60%. Такое количество холестерина значительно уменьшает размер кооперативной единицы и снижает различия между твердой и жидкой фазами.
Липидные изомеры в монослое упакованы одинаковым способом: полярные части молекул погружены в водную фазу, а углеводородные части направлены в воздух или погружены в органический растворитель.
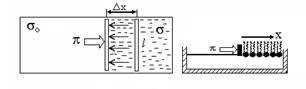
Молекулы липида в нерастворимом монослое могут двигаться вдоль поверхности воды, при этом они ударяются о подвижный барьер, создавая поверхностное давление π. Работа внешней силы против этого давления при перемещении барьера длиной l на расстоянии dx равно

С другой стороны, изменение поверхностной энергии при замещении поверхности, занятой монослоем, на чисто водную поверхность равно

где σ0и σповерхностное натяжение воды и монослоя соответственно. Отсюда
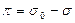
σ0 =72,8 дин/см на границе вода – воздух
≈50 дин/см на границе воды и жидкого н-алкана при 20о, а σ – зависит от вида молекул, образующих мономолекулярную пленку.
Изменения с подвижным барьером (весы Ленгмюра) позволяют непосредственно определить силу поверхностного давления от площади (А), приходящуюся на одну молекулу при постоянной температуре, если известно количество липида в монослое.
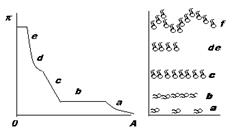
Участок а соответствует состоянию “ двумерного газа “. На участке b происходит конденсация пленки при постоянном давлении, при этом “ двумерный газ “ и “ двумерная жидкость “ сосуществуют. Пленка на участке с, характеризуется сравнительно малым коэффициентом сжатия (¶А/¶π)т называется жидкорастянутой. При дальнейшем повышении давления (участок de) происходит переход двумерной жидкости в конденсированную пленку, характеризующуюся почти полной несжимаемостью. Участок f соответствует разрушению пленки (коллажу) с образованием капель или многослойных “доменов”.
Конденсированные пленки могут быть как в жидком, так и в твердом состоянии. Жидкие конденсированные пленки удерживаются в этом состоянии только за счет внешних сил (давление барьера) и спонтанно расширяются при снятии этого давления. Наоборот, твердые (квазикристаллические) пленки удерживаются чисто внутренними силами за счет межмолекулярных взаимодействий (боковой когезии в монослое). Для таких пленок характерно сопротивление сдвигу: они не вытекают через отверстие в барьере при наличии некоторого поверхностного давления.
Кривые для реальных пленок хорошо описываются уравнением, представляющим двумерный аналог уравнения Ван-дер-Ваальса.
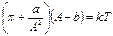
а – константа Ван-дер-Ваальса, характеризующая силу межмолекулярного взаимодействия,
b – эффективная площадь сечения молекулы (b≈А0), k – постоянная Больцмана, Т – абсолютная температура.
В зависимости от поверхностной плотности липида, он ведет себя как газ, либо жидкость, либо твердое тело.
1. Если площадь поверхности, приходящаяся в среднем на одну молекулу высшей жирной кислоты превышает 0,5 нм2, то такая степенная функция ведет себя как двумерный газ.
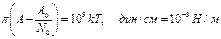
π – поверхностное давление (Н/м),
А – площадь поверхности на одну молекулу
А0 – площадь, занимаемая 1 молем
N0 – число Авогадро 6,02×1023
k – постоянная Больцмана
2. При увеличении поверхностного давления молекулы постепенно сближаются. Если площадь, приходящаяся в среднем на одну молекулу ≈0,5нм2, то монослой называется растянутым и он соответствует жидкому состоянию, имеющему другую предельную молярную площадь À0. Такому монослою соответствуют значения π – 10-3- 10-2H/м
Уравнение состояния

 πс - поверхностное давление, предельное для газообразного состояния.
πс - поверхностное давление, предельное для газообразного состояния.
3. Если поверхностное давление превышает 10-2Н/м, то монослой называется конденсированным. Все амфифильные молекулы принимают указанную выше ориентацию. Полярные головы остаются гидратированными.
Уравнение состояния

k′ - коэффициент,
а – активность растворенного вещества
Аs – площадь поверхности
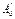 - предельная молярная площадь для конденсированного монослоя.
- предельная молярная площадь для конденсированного монослоя.
4. Дальнейшее повышение поверхностного давления приводит к потере полярными головами их гидратной оболочки с образованием твердого монослоя. При этом А (≈0,2 нм2) почти не зависит от длины углеводородной цепи, а монослой – практически несжимаем. Уравнение состояния – то же, что и для конденсированного монослоя (но с другим коэффициентом).
При еще больших поверхностных давлениях монослой разрушается и начинают образовываться мультислойные структуры.
- Понятие смещения, скорости и ускорения при вращательном движении точки
Вращательным называют такое движение твердого тела, при котором две какие-нибудь точки принадлежащие телу, остаются во все время движения неподвижными. Прямая, проходящая через эти точки, называется осью вращения. Все точки лежащие на оси так же неподвижны.
Основными кинематическими характеристиками вращательного движения твердого тела являются его угловая скорость 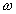 и угловое ускорение
и угловое ускорение 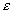 .
.
Если за промежуток времени 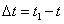 тело совершает поворот на угол
тело совершает поворот на угол 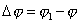 , то средняя угловая скорость будет численно равна
, то средняя угловая скорость будет численно равна
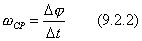
Угловой скоростью тела в данный момент t называется величина, к которой стремится средняя угловая скорость  , если
, если 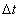 стремится к нулю.
стремится к нулю.
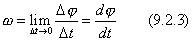
Угловая скорость твердого тела является первой производной от угла поворота по времени.
Размерность: [радиан/время]; [1/время]; [1/сек =  ].
].
Угловую скорость можно изображать вектором. Вектор угловой скорости 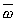 направляют по оси вращения в ту сторону, откуда вращение видно против хода часовой стрелки.
направляют по оси вращения в ту сторону, откуда вращение видно против хода часовой стрелки.
Если угловая скорость не является постоянной величиной, то вводят еще одну характеристику вращения - угловое ускорение.
Угловое ускорение характеризует изменение угловой скорости тела с течением времени.
Если за промежуток времени угловая скорость получает приращение  , то среднее угловое ускорение равно
, то среднее угловое ускорение равно
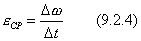
Угловым ускорением твердого тела в данный момент времени t называется величина к которой стремится 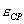 при стремящемся к нулю
при стремящемся к нулю

Как вектор, угловое ускорение 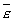 направлен так же, как и , вдоль оси.
направлен так же, как и , вдоль оси.
Если направление и совпадает, то вращение ускоренное, если противоположно, то замедленное.
Если = const, то вращение будет равномерным.
Найдем его закон. Так как  , то, интегрируя при начальных условиях t = 0,
, то, интегрируя при начальных условиях t = 0,  = 0, получаем
= 0, получаем
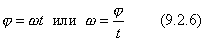
Это и есть закон равномерного вращения.
В технике вращение характеризуют оборотами в минуту n [об/мин]. Угловая скорость и обороты в минуту n связаны следующим соотношением:
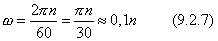
Если угловое ускорение тела все время остается постоянным, то вращение называют равнопеременным ( = const).
Найдем закон вращения, если в начальный момент t = 0, = 0 и 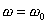 :
:
 , интегрируя получаем
, интегрируя получаем

Подставляем вместо правую часть (9.2.3), разделяем переменные и, вновь интегрируя, имеем
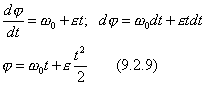
Это закон равнопеременного вращения.
Если и имеют один знак, то вращение равноускоренное. Если знаки разные - равнозамедленное. (рис. 51, а,б).
- Понятие массы и силы
Масса – мера инертности тела при поступательном движении
· Пассивная гравитационная масса показывает, с какой силой тело взаимодействует с внешними гравитационными полями — фактически эта масса положена в основу измерения массы взвешиванием в современной метрологии.
· Активная гравитационная масса показывает, какое гравитационное поле создаёт само это тело — гравитационные массы фигурируют в законе всемирного тяготения.
· Инертная масса характеризует инертность тел и фигурирует в одной из формулировок второго закона Ньютона. Если произвольная сила в инерциальной системе отсчёта одинаково ускоряет разные исходно неподвижные тела, этим телам приписывают одинаковую инертную массу.
Date: 2015-09-24; view: 662; Нарушение авторских прав