Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Основы биотехнологии 3 page
|
|
Часто некоторые клетки перевиваемых первичных клеточных культур претерпевают генетические изменения, в результате которых ускоряется их рост. Культуры клеток, которые при этом приобретают селективные преимущества, оказываются способными к неограниченному росту in vitro и называются устойчивыми клеточными линиями. Одни клеточные линии сохраняют основные биохимические свойства исходных клеток, другие нет. У большинства клеток, способных к неограниченному росту, имеются значительные хромосомные изменения, в частности отмечается увеличение числа одних хромосом и потеря других. В молекулярной биотехнологии устойчивые клеточные линии иногда используют для размножения вирусов и для выявления белков, которые кодируются клонированными последовательностями ДНК. Кроме того, они применяются для крупномасштабного производства вакцин и рекомбинантных белков.
Растительные клетки. Большой интерес представляет культура одиночных клеток. Согласно Т.А.Егоровой и соавт. (2003), ее применяют в клеточной селекции для отбора гибридных клеток и их клонирования, а также для генетических и физиологических исследований. Особенно это важно для культивирования растительных клеток. Известно, что одиночная клетка живет, но не делится в тех условиях, которые разработаны для нормального роста и размножения клеток каллусной ткани.
Напомним, что культура изолированных тканей обычно представлена каллусными и гораздо реже опухолевыми тканями. Каллусная ткань образуется в результате повреждения на целых растениях, а также в стерильной культуре на эксплантатах – фрагментах ткани или органа, используемых для получения первичного каллуса. Возникновение каллуса связано с неорганизованным делением (пролиферацией) дедифференцированных клеток. Дедифференцировка – основа создания каллусной ткани.
Культивирование одиночных клеток основано на использовании так называемого «кондиционирующего фактора» - метаболитов, выделяемых в среду культивирования делящимися клетками. Действительно, когда в питательную среду вводится одна клетка или небольшое их количество, они не делятся, так как выделяемого «кондиционирующего фактора» не хватает для индукции деления. Следовательно, необходимо повысить концентрацию этого фактора в питательной среде. Это достигается с помощью следующих методов:

Рис. 7. Использование культуры суспензионных клеток в качестве «кормящего слоя» для выращивания изолированных протопластов и одиночных клеток кукурузы (Егорова Т.А. и соавт., 2003): 1 – колонии клеток; 2 – фильтровальная бумага; 3 – алюминиевая сетка; 4 – пенополиуретан; 5 – суспензия клеток.
1. Метод ткани-«няньки» - кондиционирующий фактор выделяется находящимися рядом с одиночной клеткой кусочками ткани-«няньки».
2. Метод «кормящего слоя» - кондиционирующий фактор выделяют активно делящиеся клетки суспензионной культуры того же вида растений, что и одиночная клетка (рисунок 7).
3. Кондиционирование среды – осуществляется путем добавления в нее питательной среды, отфильтрованной от интенсивно делящихся клеток.
4. Метод культивирования одиночных клеток - осуществляется в микрокапле (объем около 20 мкл) богатой питательной среды.
Кондиционирующий фактор водорастворим, термостабилен, не заменяется фитогормонами, включает низкомолекулярные вещества. Химическая природа кондиционирующего фактора видна из следующего эксперимента. Если разделить одиночные клетки и ткань-«няньку» стеклянной пластиной, то деление клеток не наступает. Если вместо пластин поместить целлофан, то хотя и с опозданием начинается деление одиночных клеток (рисунок 8).
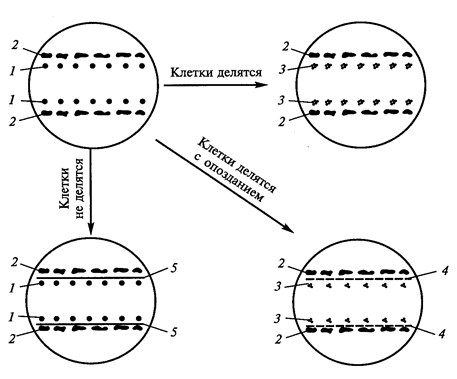
Рис. 8. Доказательство химической природы фактора кондиционирования (Егорова Т.А. и сотр., 2003): 1 – одиночные клетки; 2 – ткань-«нянька»; 3 – делящиеся клетки; 4 – целлофан; 5 – стеклянные пластинки.
Культуры клеток и тканей растений нашли широкое применение в биотехнологии: массовое клональное микроразмножение плодовоовощных и декоративных растений, их оздоровление от вирусных и других инфекций, синтез вторичных метаболитов. Известно около 20000 синтезируемых растениями вторичных метаболитов, используемых человеком: компоненты пищи, лекарства, красители, эфирные масла и др. Способность интактных растений синтезировать различные соединения обосновала предположение о возможности получение таких веществ с помощью культур клеток, тканей и органов. Это направление оказалось весьма перспективным. Синтез вторичных метаболитов происходит главным образом в суспензионной культуре клеток, в регулируемых условиях. Поэтому он не зависит от климатических факторов, от повреждения насекомыми. Культуры выращивают на малых площадях. Культуры клеток растений могут синтезировать практически все классы соединений вторичного обмена, причем довольно часто в количествах, в несколько раз превышающих их синтез в целых растениях. В культурах клеток может начаться синтез веществ, не характерных для исходного растения, либо расширяется набор синтезируемых соединений. Важная особенность культивируемой популяции клеток – ее стабильность в отношении синтеза и накопления продуктов вторичного синтеза. Синтез вторичных метаболитов в культивируемых клетках связан с внутриклеточными органеллами (тблица 2).
Таблица 2 - Внутриклеточная локализация синтеза и накопления вторичных метаболитов (Бутенко Р.Г., 1999)
| Внутриклеточные метаболиты | Синтез | Накопление |
| Алкалоиды | Пластиды, цитоплазма | Вакуоль, хлоропласты, СП |
| Монотерпены Тритерпены | Лейкопласты Хлоропласты, лейкопласты | СП Вакуоль, СП, цитозоль |
| Флавоноиды Танины Кумарины Оксикоричные кислоты | Хлоропласты Вакуоль, пластиды Вакуоль, хлоропласты ЭПР, хлоропласты, МХ | Вакуоль, хлоропласты Вакуоль, СП, ЭПР Вакуоль Вакуоль, хлоропласты |
| Цианогенные гликозиды | ЭПР | Вакуоль |
| Глюкозинолаты | ЭПР | Вакуоль |
| Бетаины | Цитозоль | Вакуоль |
Примечание: СП – свободное пространство клетки, ЭПР – эндоплазматический ретикулум, МХ – митохондрии.
Технология рекомбинантных ДНК (молекулярное клонирование или генетическая инженерия) – это совокупность экспериментальных процедур, позволяющая осуществлять перенос генетического материала (ДНК) из одного организма в другой. Рассмотрим типичную процедуру.
1. Из организма – донора нужных генов – экстрагируют нативную ДНК (клонируемая ДНК, встраиваемая ДНК, ДНК-мишень, чужеродная ДНК). Эту ДНК подвергают ферментативному гидролизу и соединяют с другой ДНК (вектор для клонирования, клонирующий вектор) с образованием новой, рекомбинантной молекулы. Образуется новая рекомбинантная молекула: конструкция «клонирующий вектор – встроенная ДНК».
2. Эту конструкцию вводят в клетку-хозяина (реципиент), где она реплицируется и передается по наследству при каждом делении. Этот процесс называется трансформацией.
3. Отбирают клетки, несущие рекомбинантную ДНК (трансформированные клетки).
4. Получают специфический белковый продукт, синтезированный клетками-хозяевами, что служит подтверждением клонирования искомого гена.
Успех молекулярного клонирования зависит:
1) от возможности воспроизводимо разрезать молекулу ДНК на фрагменты определенного размера;
2) способа их целенаправленного транспорта;
3) высокоспецифичного отбора клеток с клонированным геном и возможности индукции его экспрессии.
Для точного разрезания молекул ДНК используют высокоспецифичные ферменты рестриктазы - бактериальные рестрицирующие эндонуклеазы типа II. Например, одна из первых рестрицирующих эндонуклеаз II типа была выделена из бактерии E. coli и получила название Eco RI. Этот фермент узнает участок ДНК, содержащий специфическую палиндромную последовательность (перевертыш типа «кок», «шалаш») G-A-A-T-T-C, и катализирует разрыв между остатками гуанина (G) и аденина (А) в каждой цепи.
↓
Р-цепь-Г-А-А-Т-Т-Ц-цепь-ОН
НОцепь-Ц-Т-Т-А-А-Г-цепь-Р →
↑
Р-цепь-Г-ОН + Р-цепь-А-А-Т-Т-Ц-цепь-ОН
НОцепь-Ц-Т-Т-А-А-цепь-Р НО-Г-цепь-Р
Происходит расщепление связи между атомом кислорода при 3′-атоме углерода дезоксирибозного остатка одного нуклеотида и фосфатной группой, присоединенной к 5′-углеродному атому дезоксирибозы соседнего нуклеотида. Разрывы в цепи ДНК располагаются наискось друг от друга, в результате чего образуются одноцепочечные комплементарные концы с «хвостами» из четырех нуклеотидов в каждом (липкие концы). Каждый одноцепочечный «хвост» заканчивается 5′-фосфатной группой, а 3′-гидроксильная группа противоположной цепи как бы утоплена. К настоящему времени получены тысячи рестриктаз; их номенклатура: род микроорганизма обозначается прописной буквой, а вид – двумя строчными, штамм обычно не указыается; римские цифры – порядковый номер данной эндонуклеазы в ряду прочих рестриктаз, выделенных из данного микроорганизма (см. выше Eco RI). Палиндромные последовательности, которые атакуют рестриктазы называют сайтами узнавания. Рестриктазы существуют двух типов: 1) катализируют косое расщепление двойной спирали ДНК с образованием «липких концов» и 2) катализируют перпендикулярный разрыв с образованием «тупых концов».
Приведем примеры наиболее часто используемых ферментов рестрикции (таблица 3).
Таблица 3 - Нуклеотидные последовательности, распознаваемые типичными ферментами рестрикции (по Б.Глик и Дж. Пастернак 2002)
| Фермент | Сайт узнавания | Место разрыва | Характер концов |
| EcoRI | ГААТТЦ | Г-А | Выступающие концы |
| BamHI | ГГАТЦЦ | Г-Г | Выступающие концы |
| Pstl | ЦТГЦАГ | А-Г | Выступающие концы |
| Sau3AI | ГАТЦ | Х-Г | Выступающие концы |
| PvuII | ЦАГЦТГ | Г-Ц | Тупые концы |
| HpaI | ГТТААЦ | Т-А | Тупые концы |
| HaeIII | ГГЦЦ | Г-Ц | Тупые концы |
| NotI | ГЦГГЦЦГЦ | Г-Ц | Выступающие концы |
Рестриктазы используют для 1) изучения последовательности нуклеотидов в ДНК и 2) в генетической инженерии. В последнем случае рекомбинантные ДНК получают, когда два разных образца ДНК обрабатывают одной и той же рестриктазой с образованием фрагментов с липкими концами, а затем смешивают эти образцы. Благодаря комплементарному спариванию липких концов фрагментов разных образцов ДНК могут образовываться новые комбинации генов.
Этот ключевой механизм молекулярного клонирования поддерживается тремя способами:
1. Фрагменты ДНК, образующие комплементарные дуплексы в области липких концов удерживаются водородными связями недостаточно прочно, чтобы молекулы в растворе оставались стабильными длительное время. Для стабилизации используют фермент ДНК-лигаза бактериофага Т4, который катализирует образование фосфодиэфирных связей между концами полинуклеотидных цепей, которые уже удерживаются вместе благодаря спариванию липких концов. Этот же фермент способен «сшивать» тупые концы, которые сближаются друг с другом после связывания с ферментом.
2. Объединение разных молекул ДНК (образование рекомбинантных ДНК) требует их реплицирования в клетке-хозяине. Следовательно, одна часть рекомбинантной молекулы ДНК должна нести нужный ген, который предполагается клонировать, а другая – должна содержать информацию, необходимую для репликации в клетке рекомбинантной ДНК (клонирующие векторы).
3. При рестрикции ДНК образуется смесь разнообразных фрагментов, а затем после их лигирования с векторной ДНК образуется множество различных комбинаций. Поэтому существуют специальные системы скрининга для распознавания реципиентных клеток, содержащих ДНК с нужной для клонирования нуклеотидной последовательностью.
Для целенаправленного транспорта выделенного гена в клетку используют плазмидные векторы. Плазмиды – это внехромосомные автономно реплицирующиеся двухцепочечные кольцевые молекулы ДНК, содержащие от 1 до 200 т.п.н. и более. Плазмиды встречаются практически у всех бактерий. Их делят на высококопийные (в клетке 10-100 копий) и низкокопийные (в клетке 1-4 копии). На долю плазмидной ДНК приходится 0,1-5,0% суммарной клеточной ДНК. Классификация плазмид по типу содержащейся информации:
- F-плазмиды содержат информацию для собственного переноса из одной клетки в другую;
- R-плазмиды несут гены устойчивости к антибиотикам;
- плазмиды деградации содержат гены, ответственные за утилизацию необычных метаболитов;
- криптические плазмиды несут гены, выполняющие скрытые (латентные) функции;
- плазмиды с узким спектром хозяев несут специфический сайт начала (инициации) репликации и могут реплицироваться только в клетках одного вида;
- плазмиды с широким спектром хозяев – содержат неспецифичный сайт инициации репликации, что позволяет им реплицироваться в разных бактериальных клетках.
Для целей генетической инженерии требуются плазмиды, обладающие тремя важными свойствами:
1) небольшой размер (менее 15 т.п.н.) необходимый для эффективного переноса;
2) наличие уникального сайта рестрикции, в который может быть осуществлена вставка;
3) наличие одного или более селективных генетических маркеров для идентификации реципиентных клеток, несущих рекомбинантную ДНК.
Природные (не модифицированные) плазмиды обычно лишены одного или нескольких свойств, необходимых для эффективного переноса, поэтому качественные плазмидные векторы приходится также создавать методами генетической инженерии.
В технологии рекомбинантных ДНК классическим является плазмидный вектор рВR322. Номенклатура: р – от английского plasmid, следующие буквы имеют отношение к описанию вектора или истории его создания (для данного вектора буквы BR указывают на авторство Ф. Боливара и Р. Родригеса); цифра 322 номер из исследовательского протокола. Размер плазмиды рВR322 4361 п.н. Эта плазмида имеет:
- два гена устойчивости к антибиотикам ампициллину (AmpR) и тетрациклину (TetR);
- уникальные сайты узнавания ферментами рестрикции для Bam H1, Hind III, Sal I в гене TetR, один Pst I-сайт в гене AmpR (разрыв генов TetR и AmpR и встраивание в это место клонируемого гена лишит трансформированную плазмидой клетку устойчивости к соответствующему поврежденному гену антибиотику –тетрациклину или ампицициллину);
- один сайт для Eco RI, находящийся за пределами кодирующих последовательностей;
- сигнал начала репликации, обеспечивающий репликацию исключительно в бактериальных клетках кишечной палочки.
Принцип работы плазмидного вектора рВR322:
1. Плазмиду рВR322 обрабатывают рестриктазой, которая катализирует ее разрыв в единственном сайте, расположенном в одном из генов устойчивости к тому или иному антибиотику. Образуется линейная молекула с липкими концами.
2. Донорную ДНК, содержащую нужный ген, обрабатывают той же рестриктазой, что приводит к образованию фрагментов ДНК с липкими концами.
3. Линейные молекулы плазмиды рВR322 смешивают с обработанной донорной ДНК. Поскольку липкие концы этих двух ДНК взаимно комплементарны, они спариваются с образованием гибридных молекул.
4. Для стабилизации гибридных молекул ДНК производят обработку ДНК-лигазой фага Т4 в присутствии АТФ. В результате этого образуется множество разных комбинаций фрагментов. Кроме того, образуются нежелательные продукты, объединяющие между собой фрагменты донорной ДНК и исходные плазмидные ДНК. Для их уменьшения обрабатывают рестрицированную плазмидную ДНК щелочной фосфатазой, отщепляющей от линейной молекулы 5′-фосфатные группы. В результате ДНК-лигаза не сможет сшить концы дефосфорилированной линейной плазмидной ДНК. Что касается собственно рекомбинантных молекул ДНК, то два из четырех одноцепочечных разрыва при действии ДНК-лигазы устраняются, и конструкция оказывается стабильной благодаря образовавшимся фосфодиэфирным связям.
5. После введения гибридной ДНК в клетку хозяина происходит ее репликация и образуются новые кольцевые молекулы уже без разрывов.
Трансформация и отбор. Введение рекомбинантной ДНК в клетку-хозяина называется трансформацией. Для ее осуществления используют специально разработанные приемы, например, подвергают клетки высокотемпературному воздействию и обрабатывают их хлоридом кальция. Однако эффективность трансформации остается низкой. Положительный результат получается примерно в одной клетке из тысячи (введение в клетку плазмиды со встроенным фрагментом чужеродной ДНК – гибридная плазмида). Проникновение в клетку экзогенной ДНК еще не означает, что она будет поддерживаться в клетке-хозяине. Для сохранения рекомбинантной ДНК в клетке-хозяине в первоначальном виде необходимо отсутствие генов, кодирующих синтез эндогенных рестриктаз, разрушающих ДНК. Необходимо также, чтобы клетка имела фенотип RecA- (такие клетки неспособны к общей рекомбинации).
Следующий этап – это идентификация клеток, содержащих рекомбинантную ДНК. При использовании плазмиды рВR322, в которой чужеродная ДНК встраивается в сайт Bam HI, специфическая идентификация состоит из двух этапов:
1. Клетки после трансформации высевают на питательную среду, содержащую ампицилин. Вырастают только клетки с наличием интактного гена AmpR (или в составе интактной плазмиды рВR322, или в составе гибридной плазмиды). Нетрансформированные клетки чувствительны к ампициллину. Сайт Bam HI расположен в гене TetR плазмиды рВR322; встраивание в этот ген фрагмента ДНК прерывает кодирующую последовательность, в результате чего устойчивость к тетрациклину утрачивается. Следовательно, клетки, которые несут гибридную плазмиду, устойчивы к ампициллину, но чувствительны к тетрациклину, а клетки, получившие интактную плазмиду рВR322, несут ген TetR и устойчивы как к ампициллину, так и к тетрациклину.
2. Клетки, выросшие на среде с ампициллином, переносят на среду с тетрациклином. Клетки, образующие колонии на чашках с тетрациклином, содержат интактную плазмиду рВR322 (они устойчивы к обоим антибиотикам). Клетки, не выросшие на среде с тетрациклином, чувствительны к этому антибиотику, а значит они содержат гибридную плазмиду рВR322. Среди колоний, выросших на среде с ампициллином, выделяют те, которые оказались чувствительны к тетрациклину, и из каждой колонии получают индивидуальные клеточные клоны. Чаще объединяют все колонии, устойчивые к ампициллину и чувствительные к тетрациклину, культивируют их вместе. Далее проводят дополнительный скрининг и идентифицируют те клетки, которые несут гибридную плазмиду рВR322 со специфической вставкой.
В настоящее время известно много плазмидных векторов. Например, на основе плазмиды pUC создан вектор, содержащий ген, который кодирует убивающий клетку белок. Этот остроумный подход позволяет облегчить сортировку и выделение трансформированных клеток.
Итак, системы клонирования должны включать 1) несколько сайтов для специфической рестрикции; 2) удачную структуру вектора, транспортирующего клонируемый ген; 3) достаточно простую идентификацию клеток с рекомбинантными ДНК. Уникальные сайты рестрикции выполняют в опытах с рекомбинантной ДНК двойную функцию: 1) обеспечивают встраивание чужеродной ДНК в вектор; 2) обеспечивают вырезание клонированной последовательности из вектора. Для рутинных процедур молекулярного клонирования используют E.coli, Bacillus subtilis, Agrobacterium tumefaciens. Во многих случаях в векторы, которые функционируют в E.coli, можно встроить второй сайт инициации репликации, обеспечивающий их репликацию в других клетках. Это так называемые челночные векторы позволяют сначала проводить клонирование в E.coli, а затем внедрять готовую конструкцию в другие клетки.
Геномная библиотека. Процесс разделения геномной ДНК на клонируемые элементы и введение этих элементов в клетки-хозяева называется созданием геномной библиотеки (банка клонов, банка генов).
Для молекулярной биотехнологии важна идентификация структурных генов, кодирующих определенные белки. У прокариот структурные гены непрерывны, а у эукариот кодирующие участки генов – экзоны разделены некодирующими – интронами. Поэтому технологии клонирования генов у прокариот и эукариот имеют различия.
У прокариот нужная последовательность для клонирования (ДНК-мишень) часто не превышает 0,02% от суммарной хромосомной ДНК. Технология клонирования и отбора ДНК-мишени включает: 1) гидролиз суммарной ДНК рестриктазой; 2) каждый из получившихся фрагментов встраивают в вектор; 3) обнаружение специфической клеточной линии (клон), которая несет нужную последовательность; 4) выделение и характеристика этой клеточной линии. У эукариот (прерывистые гены) вначале выделяют иРНК, затем с помощью обратной транскриптазы синтезируют кДНК (ДНК, комплементарная иРНК) и уже ее используют для молекулярного клонирования и создания геномной библиотеки.
Следующий после создания геномной библиотеки этап – это поиск клона (клонов), несущего искомую последовательность ДНК. Для этого используют три метода.
1. Гибридизация с меченым ДНК-зондом с последующим радиоавтографическим анализом. После трансформации высевают клетки на чашки с питательной средой и переносят выросшие колонии на твердую подложку (нитроцеллюлозный или нейлоновый фильтр). Лизируют клетки. Производят депротеинизацию и денатурацию ДНК. Фиксируют ДНК на подложке. Наносят на фильтр меченый зонд и производят отжиг, а затем радиоавтографию. Колонии на исходной чашке, которые содержат гибридизовавшуюся ДНК, выделяют и культивируют.
2. Иммунологический скрининг. В результате экспрессии клонированного гена образуется белок, который можно обнаружить иммунологическими методами. Для этого все клеточные линии (клоны) библиотеки высевают на чашки с питательной средой. Выросшие колонии переносят на фильтр, клетки лизируют, а высвободившиеся белки фиксируют на фильтре. Затем на фильтр наносят первые антитела, которые специфически связываются с данным белком (антигеном). Все несвязавшиеся антитела удаляют, а фильтр помещают в раствор вторых антител, специфичных в отношении первых антител. Во многих тест-системах используют конъюгаты вторых антител с ферментом (например, щелочной фосфатазой). После отмывания фильтра добавляют бесцветный субстрат. Если вторые антитела связываются с первыми, то под действием фермента происходит гидролиз субстрата с образованием окрашенного вещества в том месте, где идет реакция.
3. Скрининг по активности белка. Если искомый ген кодирует фермент, не синтезируемый клеткой-хозяином, то для обнаружения клонов используют цветную реакцию, катализируемую данным ферментом.
Клонирование крупных фрагментов ДНК. Чтобы иметь возможность клонировать целый ген, донорную ДНК расщепляют лишь частично. При этом получаются фрагменты разной длины, из которых затем создают геномную библиотеку. Для клонирования крупных фрагментов ДНК были сконструированы векторы на основе бактериофагов лямбда и Р1, а также плазмиды F.
В заключение этого раздела пособия следует отметить, что конечный результат молекулярного клонирования ДНК лишь тогда положителен, если будет доказано получение нативного структурного гена.
Для целей молекулярного клонирования используются химический синтез, определение нуклеотидной последовательности и амплификация ДНК.
Химический синтез ДНК используют для получения 1) одноцепочечных цепей дезоксирибонуклеотидов (олигонуклеотиды) и 2) двухцепочечных цепей ДНК (синтез генов).
Синтез олигонуклеотидов длиной около 50 нуклеотидов осуществляется с помощью ДНК-синтезаторов с использованием фосфорамидитного метода. В отличие от биологического синтеза ДНК (присоединение каждого последующего нуклеотида по 3′-ОН группе дезоксирибозы, в химическом синтезе каждый новый нуклеотид присоединяется к 5′-ОН группе, т.е. 5′-концу олигонуклеотида). Синтезированные олигонуклеотиды используются:
- для конструирования фрагментов или целых генов;
- для амплификации (усиление синтеза) специфических фрагментов ДНК;
- для направленных мутаций изолированных ДНК;
- в качестве зондов при гибридизации (нуклеотидную последовательность зондов длиной 20-40 звеньев находят из данных об аминокислотной последовательности соответствующих белков);
- в качестве линкеров, облегчающих клонирование (линкер – синтетический олигонуклеотид, содержащий сайт рестрикции; используется для соединения векторной и клонируемой ДНК, к концам которой по методу сшивания тупых концов присоединены линкеры);
- олигонуклеотиды длиной 17-24 звеньев служат праймерами при секвенировании ДНК и проведения полимеразной цепной реакции (ПЦР).
Синтез генов из химически синтезированных отдельных комплементарных цепей олигонуклеотидов. Как правило, синтетические двухцепочечные гены собирают из модулей – одноцепочечных фрагментов длиной от 20 до 60 нуклеотидов с перекрывающимися концами. Нуклеотидные последовательности цепей задают так, чтобы после отжига (образование комплементарных двухцепочечных фрагментов) концевые сегменты гена имели тупые концы. Каждый внутренний сегмент имеет выступающие 3′- и 5′-концы, комплементарные таковым соседнего сегмента. После сборки гена остается сшить одноцепочечные разрывы с помощью ДНК-лигазы Т4. Синтетические гены могут быть сконструированы так, чтобы помимо белок-кодирующих последовательностей они содержали концевые участки, обеспечивающие:
- встраивание в клонирующий вектор (сайты для рестриктаз);
- сигнальные последовательности для правильной инициации и терминации транскрипции и трансляции..
Секвенирование ДНК – это определение нуклеотидной последовательности полинуклеотидных цепей. Без знания нуклеотидной последовательности генов невозможно осуществлять молекулярное клонирование. Для секвенирования используют несколько методов.
1. Дидезоксинуклеотидный метод. Дидезоксинуклеотид – это полученный искусственным путем нуклеотид, лишенный 2′- и 3′-гидроксильных групп при углеродных атомах сахара. У дезоксирибонуклеотидов, входящих в ДНК, отсутствует только 2′-гидроксильная группа. Как указано выше, удлинение цепи во время репликации ДНК происходит в результате присоединения очередного нуклеозидтрифосфата к 3′-гидроксильной группе последнего нуклеотида растущей цепи. Если же присоединяемым нуклеотидом является дидезоксинуклеотид, то синтез ДНК останавливается, поскольку следующий нуклеотид не может образовать фосфодиэфирную связь. Остановка синтеза ДНК – ключевой этап дидезокси-метода. Для секвенирования в разных пробирках одновременно проводят четыре реакции синтеза ДНК, каждая – в присутствии одного из четырех дидезоксинуклеотидов (ддАТФ, ддГТФ, ддЦТФ и ддТТФ). Продукты реакций разделяют с помощью гель-электрофореза, проводят радиоавтографию и «считывают» с радиоавтографа нуклеотидную последовательность синтезированного фрагмента ДНК.
2. Для секвенирования более длинных генов используют систему на основе фага М13. В ДНК фага встраивают фрагмент ДНК длиной до 500 нуклеотидов, который хотят секвенировать. Эту рекомбинантную ДНК легко получить в одноцепочечной форме и использовать ее в качестве матрицы для секвенирования вставки. Можно использовать также двухцепочечные плазмиды, содержащие клонированную ДНК.
3. Для определения нуклеотидной последовательности протяженных клонированных сегментов сначала подбирают синтетический олигонуклеотидный праймер, комплементарный участку, соседствующему со вставкой. Затем с помощью дидезокси-метода секвенируют первые 250-300 нуклеотидов. Затем по результатам секвенирования синтезируют второй праймер и определяют последовательность следующих 250-350 нуклеотидов клонированного участка и т.д. Этот метод, называемый «праймер-опосредованной прогулкой» (или «блуждающей затравкой»), позволяет секвенировать протяженные фрагменты ДНК без их субклонирования, как в случае системы на основе фага М13.
В таблице 4 представлен прогресс в секвенировании генов.
Таблица 4 - Историческая справка о секвенировании ДНК генов (Primrose S.B., Twyman R.M., 2003)
| Секвенированный ген | Год | Размер гена | Комментарий |
| Бактериофаг фх174 | 5,38 kb | Первый ген | |
| Плазмида рBR 322 | 4,30 kb | Первая плазмида | |
| Бактериофаг А | 48,5 kb | ||
| Вирус Эпштейн-Барра | 172 kb | ||
| Дрожжевая хромосома III | 315 kb | Первая хромосома | |
| Haemophylus influence | 1,8 Мb | Первый геном организма-клетки | |
| Saccharomyces cerevisiae | 12 Mb | Первый геном клетки-эукариота | |
| Ceanorhabditis elegans | 97 Mb | Первый геном мультиклеточного организма | |
| Drosophila melanogaster | 165 Mb | ||
| Arabidopsis thaliana | 125 Mb | Первый геном растения | |
| Homo sapiens | 3000 Mb | Первый геном млекопитающего | |
| Pufferfish (Fugu rubripes) | 400 Mb | Наименьший геном позвоночных | |
| Мышь (Mus musculis) | 2002/3 | 2700 Mb | Генетическая модель человека |
Date: 2015-09-27; view: 1392; Нарушение авторских прав