Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Эколого-биологические последствия Чернобыльской катастрофы 5 page
|
|
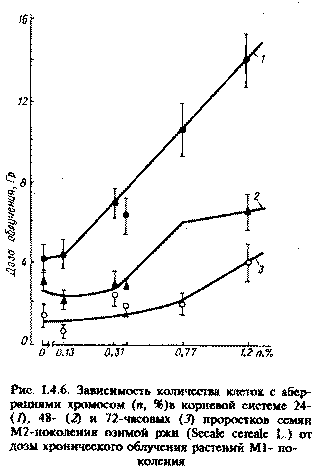
Рис. 1.4.6. Зависимость количества клеток с аберрациями хромосом (n, %) в корневой системе 24-(1), 48- (2) и 72-часовых (3) проростков семян М2-поколения озимой ржи (Secale cereale L.) от дозы хронического облучения растений М1-поколения
Кроме того, было установлено [126] увеличение количества клеток с аберрациями хромосом и хроматид в корневой меристеме проростков скерды кровельной (Crepis tectorum L.), выращенных из семян растений, собранных на территории зоны ЧАЭС при условиях гамма-фона 5-10 мР/ч в первый и 0,02 - 20 мР/ч во второй годы после аварии. В проростках первого года максимальная частота клеток с аберрациями достигала 10,2 - 15,3 %, причем нередко наблюдались клетки со множественными аберрациями. Во второй год на участках с максимальным уровнем фона ьыло обнаружено 1,4 - 2,2% клеток с аберрациями хромосом. При минимальных мощностях облучения частота клеток с аберрациями составила всего 0,3 - 0,5%; эти значения соответствуют спонтанному уровню. Кроме того, отмечали также появление растений с измененным кариотипом, что свидетельствует об активных микроэволюционных процессах в хронически облучаемых популяциях.
Имеются данные [88], свидетельствующие о повышении количества стерильной пыльцы у растений, растущих на территориях, загрязненных радионуклидами. Так, исследуя две популяции фиалки утренней (Viola matutina klok), произрастающей на двух участках 30-километровой зоны ЧАЭС, различающихся на 3 порядка по величинам поглощенных доз, выявили в 1987 и 1988 гг. увеличение в 1,5 - 2 раза количества стерильных пыльцевых зерен. При уровне мощности дозы на высоте 2 м 4 - 5 Гр/год наблюдали частичную женскую стерильность сосны (Pinus sylvestris L.), выражавшуюся в снижении гамето-фитной выживаемости семяпочек, опыленных в 1986 г. и уменьшении эмбриональной выживаемости семяпочек, опыленных в 1985 г.
Безусловно, стерильность - неспособность организма образовывать гаметы или достаточное их количество, не приводит к генетическому повреждению, а лишь снижает число потомков. Но при действии ионизирующей радиации стерильность, как правило, является следствием генных или хромосомных мутаций, обусловливающих нарушение мейоза - основного звена гаметогенеза.
Прямым свидетельством генетического действия ионизирующих излучений на пыльцу является непосредственная регистрация в них мутаций. У растений, выращиваемых на почвах 30-километровой зоны ЧАЭС с общей бета-активностью 2,6·103- 6,3·104Бк/кг, в пыльце ячменя (Hordeum vulgare L.) отмечали увеличение частоты waxy-изменений в 2 - 3 раза [34].
Таблица І.4.7
Количество хлорофилльных мутаций типа альбина у ржи (Secale cereale L.) и ячменя (Hardeum vulgare L.), %
| Культура | Контроль | 1986 г. | 1987 г. | 1988 г. | 1989 г. |
| Рожь Киевская 80 | 0,01 | 0,14 | 0,40 | 0,91 | 0,71 |
| Рожь Харьковская 03 | 0,02 | 0,80 | 0,99 | 1,20 | 1,14 |
| Ячмень | 0,40 | 0,90 | 0,74 | 0,80 | 0,91 |
Примечание. Семена получены в 30-километровой зоне ЧАЭС.
В первые месяцы после аварии при выращивании растений ржи (Secale cereale L.) и ячменя (Hardeum sativum jessen) в условиях закрытого грунта на почвах, привезенных из 30-километровой зоны, было обнаружено увеличение частоты встречаемости различных типов хлорофилльных мутаций. Преобладали нежизнеспособные мутации типа альбина. В последующие годы семена, собранные в зоне, высевали вновь. У ржи и ячменя, выращиваемых в 30-километровой зоне на почве с суммарной гамма-активностью 1·105Бк/кг, регистрировали многократное превышение спонтанного уровня выхода хлорофилльных мутаций (табл. 1.4.7). При этом было отмечено, что в каждом последующем поколении мутации не элиминировались, поскольку высеянные семена подвергались последующему прессингу радионуклидов.
Выход морфологических аномалий у 4 сортов озимой пшеницы (Triticum vulgare L.) при их выращивании в течение 3 лет на почве с суммарной активностью гамма-излучения 5,2·104- 1,8·105Бк/кг в 1986 - 1987 гг. превысил 40 %, но в 1988 г. уменьшился. Спектр выявленных аномалий довольно широк: в первый год после аварии наиболее частым (до 49 %) было образование в колосьях стерильных зон - череззерница; в 1987 г. количество аномалий этого типа достигало 30 %, а в следующем поколении снижалось до 1,9 %. Часто встречались колосья с дополнительными колосками ("грыжи") и укороченные колосья. К распространенным изменениям структуры колоса следует также отнести аномалии типа скверхед. Встречались также растения с повышенной остистостью, неравномерной остистостью, колос "елка", изменения окраски стебля и другие. У разных сортов выход морфологических аномалий проявляется по-разному, что может быть связано, с одной стороны, с различной их радиочувствительностью, а с другой - с неодинаковой способностью к накоплению радионуклидов.
Безусловно, отмечаемые морфологические аномалии, отклонения от нормы, как правило, имеют место лишь в поколениях облучаемых организмов и обычно не закрепляются в потомстве. Тем не менее их стойкое проявление во все последующие годы, вплоть до 1994 г., позволяет предполагать, что многие из этих изменений носят генетический характер и свидетельствуют о том, что под влиянием ионизирующих излучений радионуклидов, выброшенных при аварии на ЧАЭС, в популяциях растений идет мутационный процесс.
4.4.6. Действие на растения инкорпорированных радионуклидов
Радиочувствительность растений к облучению, создаваемому в результате накопления в тканях радионуклидов, изучена меньше, чем радиочувствительность к облучению от внешних источников. Главным образом это обусловлено тем, что большинство видов растений обладает более высокой по сравнению с млекопитающими радиоустойчивостью, и даже при накоплении радионуклидов в значительных количествах действие излучений у них проявляется гораздо в меньшей мере. Растениям обычно отводится роль переносчиков радионуклидов по пищевой цепочке "почва - растение - животное - человек" или "почва - растение - человек".
Тем не менее при поступлении и накоплении радионуклидов в относительно радиочувствительных видах растений в зависимости от физико-химических характеристик радиоактивных веществ, их количества, мест локализации могут наблюдаться те же радиобиологические эффекты, что и при эквивалентных поглощенных дозах внешнего облучения: радиационная стимуляция, морфологические изменения, лучевая болезнь, ускорение старения и сокращение продолжительности жизни, гибель, генетическое действие. Однако поступившие внутрь растений радиоактивные вещества могут обладать повышенной по сравнению с внешним облучением опасностью, вызванной рядом причин. Первая и основная из них - способность некоторых радиоактивных веществ избирательно накапливаться в отдельных тканях и органах, что приводит к относительно высоким локальным уровням облучения. Вторая причина - увеличение опасности действия альфа-и бета-излучателей, которые мало влияют на растения в условиях внешнего облучения, но могут стать необычайно сильным источником ионизирующей радиации при поступлении внутрь клеток и клеточных структур. Третьей причиной являются, как правило, длительные сроки облучения. Ввиду специфики метаболизма растений радионуклиды, кумулируясь в тканях, практически не выводятся из них в отличие от животных. При огромных периодах полураспада90Sr,137Cs,239Pu даже у многолетних растений эти радионуклиды будут обусловливать облучение в течение всего онтогенеза.
И если в первый год аварии облучение растений было обусловлено в значительной степени за счет внешнего облучения, то в последующие годы с начавшейся миграцией радионуклидов по трофической цепочке, оно почти исключительно обусловливается внутренним облучением. Но если у одно-двухлетних растений за сравнительно короткий период вегетации не всегда успевают сформироваться достаточно высокие дозы облучения, то у многолетних растений в течение десятков лет могут сложиться сравнительно высокие уровни облучения. Более того, подавляющее количество видов многолетних растений, как правило, чувствительнее к действию радиации, чем одно-двухлетние травянистые виды.
В зоне Чернобыльской катастрофы в кроне древесных растений первоначально было задержано от 60 до 90 % выпавших на лес радионуклидов. После выпадения радиоактивных осадков на лесную растительность начинается их вертикальная миграция под влиянием сил гравитации, атмосферных осадков, движения воздуха, с листовым опалом. в результате чего радионуклиды перемещаются в нижние слои крон, под полог леса. Скорость вертикальной миграции зависит от физико-химических характеристик радиоактивных выпадений, типа и возраста древостоя, метрологических условий, времени года. Спустя некоторый период, который в хвойных лесах может измеряться несколькими годами, основная масса радиоактивных веществ переходит в лесную подстилку и в верхний слой почвы. Именно из него через 4 - 5 лет в лиственном лесу и через 8 - 10 лет в хвойном (что обусловлено особенностями в скорости вертикальной миграции радионуклидов и более быстрой минерализацией листьев по сравнению с хвоей) начинается активное поступление радионуклидов в древесные растения через корни.
Если механизмы поступления и усвоения радионуклидов однодвухлетними травянистыми и многолетними древесными растениями в основном не различаются, то характер из накопления имеет принципиальные различия. Многолетние древесные способны к существенному накоплению радионуклидов во всех органах - листьях, ветках, коре, древесине. Многолетний замкнутый цикл радионуклидов по цепочке "листья - лесная подстилка - почва - корни - ствол - листья" и так далее может приводить к существенному радиоактивному загрязнению всех органов. У одного из самых радиочувствительных видов - сосны - это может приводить к самым различным типам нарушений. Повреждаться могут и другие виды. Ель, например, обладает более высокой радиоустойчивостью, но она является кальциефилом и способна накапливать -Sr в больших количествах, чем сосна и другие виды.
Различные радионуклиды не только по-разному накапливаются отдельными видами растений, но и по-разному концентрируются в различных органах, приводя к различной степени облучения. На радиоавтографах целых растений, полученных при введении различных радионуклидов, как правило, бывает хорошо определена неравномерность их распределения по отдельным органам. Четко прослеживаются места концентрации, сосредоточенные, как правило, в тканях, которые обладают высокой метаболической активностью (меристема и генеративные органы) и наиболее высокой радиочувствительностью. Именно поэтому как при внешнем, так и при внутреннем облучении радиобиологические эффекты, в первую очередь, проявляются на этих группах клеток. В частности, в меристемах наблюдаются замедление или ускорение деления клеток, формирование различного рода морфологических изменений, замедление роста органов и растения в целом, гибель меристем и др.
В год аварии в пяти областях Украины (Киевской, Черниговской, Житомирской, Черкасской и Винницкой), в том числе в различных местах 30-километровой зоны, были собраны образцы семян 9 сортов озимой пшеницы (Tviticum vulgare L.), растения которой в течение 3 мес - с фазы кущения (время аварии) до созревания - подвергались облучению радиоактивными выбросами. Максимальный уровень суммарной бета- и гамма-активности семян достигал 3,7·104Бк/кг. По общепринятым методикам экспериментального мутагенеза в последующих б поколениях изучались частоты и спектр мутационной изменчивости растений. У 18,4 % образцов выявили достоверное увеличение (более чем в 2,7 раза) частоты хромосомных аберраций в клетках меристемы первичного корешка прорастающих семян. В одном случае у проростков из семян растений, выросших в 16 км от станции, наблюдали увеличение в 32 раза количества аберраций. Регистрировали увеличение в 2,7 - 8,7 раза частоты видимых мутаций. Их спектр охватывал 27 типов мутаций по признакам структуры стебля, листа и колоса, физиологическим признакам. Спецификой спектра были сравнительно частые мутации по генам карликовости, а также редко встречающиеся в обычных условиях мутации типа "ощипанный колос" и "разветвленный колос". Мутации полимерных генов по элементам структуры урожая в последующих поколениях привели к изменению продуктивности мутантов и в итоге к потере типичности сортов.
При выращивании проростков гороха (Pisum sativum L.) и кукурузы (Zea mays L.) на водных вытяжках почв из 10-километровой зоны ЧАЭС, имеющих суммарную бета-активность 5·10-9- 5·10-8Ки/л, наблюдали [46] заметное снижение относительной скорости роста растений.
Радиационную стимуляцию, выражавшуюся в ускорении прорастания семян, роста растений, прохождении фаз развития, повышении фотохимической активности хлоропластов, увеличении содержания хлорофилла, наблюдали у желтого люпина (Lupinus luteus L.), выращенного из семян, которые были собраны в 30-километровой зоне ЧАЭС при уровне загрязнения по137Cs Ки/км2. Такие семена содержали137Cs 2,31·103и90Sr 1,65·102Бк/кг [115].
Наибольшую чувствительность к радиоактивному загрязнению проявили сосновые леса по причине высокой радиочувтвительности этого вида растений, значительной задерживающей способности хвои и, как следствие, высокого вклада бета-излучения в дозовую нагрузку [49]. Общая поглощенная доза в хвое и почках примерно в 10 раз больше дозы внешнего гамма-облучения. Именно поэтому на участках с высоким уровнем загрязнения в активно растущих меристемах обнаруживали наиболее сильные повреждения, приводящие к их массовой гибели, пролифирации боковых почек с неупорядоченной ориентацией, образованию укороченных побегов с мелкой или гигантской хвоей, искривлению иголок хвои, нарушению ориентации молодых вегетативных побегов. К концу лета 1986 г. проявились летальные эффекты облучения сосны. В течение последующего осенне-зимнего периода было выявлено, что общая площадь погибшего лесного массива, примыкающего с запада к промплощадке АЭС, достигает 500 га.
Спустя 4 года после аварии наблюдалось снижение в 5 - 10 раз продуктивности семян у аборигенной популяции подорожника ланце-толистного (Plantago lanceolata L.), произрастающей в местах с уровнем гамма-фона от 2,8 до 1334 пА/кг [120]. По-видимому, данный тип радиационного поражения растений можно связать с угнетающим действием многолетнего хронического облучения на популяцию этого радиочувствительного представителя травяного покрова, не выделяя вклад внешнего и внутреннего облучения. Кроме того, без сомнения, можно утверждать, что на 4-й год после аварии, когда внешнее облучение по сравнению с первоначальным поставарийным периодом уменьшилось во много раз, фактор внутреннего облучения в радиационном повреждении растений играет определяющее значение. Особенно это касается травянистых растений с мочковатой корневой системой, формирующих основную массу корней в верхнем, наиболее загрязненном радионуклидами слое почвы.
Тем не менее следует признать, что главная опасность накопления растениями радионуклидов состоит не в их традиционной угрозе для растений, а в том, что они являются главным звеном миграционной цепочки на пути передачи радионуклидов более радиочувствительным видам живых организмов - млекопитающим и, в первую очередь, человеку. 4.5. Влияние аварии на биоценоз
4.5.1. Действующие факторы
Чтобы правильно понять причины изменений, наблюдавшихся в популяциях растений и животных после Чернобыльской катастрофы, надо принимать во внимание не только действие радиации, но и многие другие факторы, прямо или косвенно связанные с аварией на ЧАЭС или не имеющие с ней никакой причинной связи.
1. Радиационное поражение растений и животных.
2. Нарушение трофических цепей, освобождение экологических ниш в результате гибели или снижения численности видов-прокормителей или видов-конкурентов.
3. Радиационная стимуляция (гормезис).
4. Химическое загрязнение среды нерадиоактивными веществами, применявшимися для ликвидации последствий аварии на ЧАЭС.
5. Сплошная инженерная дезактивация особо загрязненных лесных кварталов, лугов, населенных пунктов (захоронение, засыпка песком).
6. Подтопление и образование мелководных водоемов после перекрытия мелиоративной сети дамбами и последующий спуск этих водоемов.
7. Подъем минеральной взвеси в р.Припять в результате защитных гидротехнических работ в русле и на берегах, массовое использование комплексообразователей в защитных гидротехнических сооружениях.
8. Искусственный разгон облаков над зоной отчуждения, увеличение уровня солнечной радиации.
9. Эвакуация населения, крупного рогатого скота, отстрел домашних хищников.
Ю. Прекращение обработки пахотных земель, сбора урожая, выпаса и сенокошения, ухода за садами, применения удобрений и пестицидов.
11. Прекращение промышленных рубок, рубок ухода, расчистки сухостоя, расчистки просек в лесах зоны отчуждения.
12. Снятие рекреационной нагрузки на природные угодья, прекращение промышленного рыбоводства на пруде-охладителе ЧАЭС, промыслового и спортивного рыболовства и охоты.
13. Продолжающаяся сукцессия растительного и животного населения Киевского водохранилища после его заполнения.
14. Климатические особенности послеаварийных лет: теплая весна 1986 г., многоснежная и холодная зима 1986 - 1987 гг., мягкие зимы последующих лет, низкие уровни весенних паводков, дождливое лето 1988 и 1991 гг., засушливые весна и лето 1992 г.
15. 11-летние циклы солнечной активности, с которыми синхрони-зуется динамика численности многих видов растений и животных.
Эти и другие менее значительные факторы порождают сложные переплетающиеся цепочки событий в живой природе, которые выражаются в смене доминирующих видов, перераспределении и изменении численности видов растений, животных и микроорганизмов. Они* затрагивают все главные звенья биоценоза: первичных продуцентов биомассы, консументов разных уровней - растительноядных животных и хищников - и редуцентов, возвращающих органогенные элементы из отмершей биомассы снова в круговорот веществ. Действие радиации непосредственно проявлялось только на ограниченной территории ближней зоны вокруг станции (радиусом 5-10 км), на остальной территории зоны отчуждения (свыше 99 %) ход событий в природных угодьях определяется в основном вторичными экологическими последствиями катастрофы.
4.5.2. Радиационное поражение биоценозов
Гибель хвойных лесов. Высокая радиочувствительность хвойных растений, усиленная тем обстоятельством, что авария произошла в период распускания почек, была причиной поражения насаждений сосны обыкновенной (Finns silvestris L.) и разрушения экосистемы сосновых лесов на наиболее загрязненных территориях вблизи атомной станции [61]. Особую роль в поражении фотосинтезирующих органов и ростовых тканей сыграло бета-излучение от осевших на поверхность растений радионуклидов; вклад внешнего гамма-облучения в поглощенную дозу составил всего 10 % [114].
На основании установленной зависимости степени поражения сосны от мощности экспозиционной дозы, рассчитанных поглощенных доз и вызванных ими эффектов поражения была составлена карта (рис. 1.4.8) радиационного поражения сосновых лесов [63].
Зона летального поражения характеризуется полной гибелью сосны. Она ограничена изолинией мощности экспозиционной дозы (МЭД) по гамма-излучению 0,5 Р/ч на 01.06.86. Расчетная поглощенная доза по гамма-излучению в зоне летального поражения составила 80 - 100 Гр. Общая площадь погибшего леса 580 га.
Хвоя погибших сосен приобрела яркую коричнево-бурую окраску (так называемый рыжий лес). "Рыжий лес" простирается вдоль основных следов радиоактивного выброса на 5 - 7 км на север, 2 - 3 км на запад, 1,5 - 2 км на юг. Отдельные пятна "рыжего леса" встречаются на расстоянии до 9 км к северу и 6 км к западу от ЧАЭС.
Лиственные леса и кустарники, травяные ценозы и мохово-лишайниковый покров в зоне летального поражения испытали усыхание вершин, угнетение ростовых процессов и повреждение репродуктивных органов в течение первого сезона вегетации.
В зоне сублетального поражения (расчетная поглощенная доза 10 - 20 Гр при максимальной МЭД 0,1 - 0,5Р/ч) произошла полная гибель молодых экземпляров сосны при частичном омертвении молодых побегов взрослых деревьев. Наряду с молодняками наиболее пострадали ослабленные насаждения сосняков-беломошников на бедных песчаных почвах. Площадь сосновых лесов в этой зоне - 3750 га.
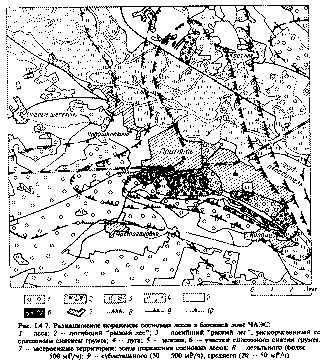
Рис. 1.4.7. Радиационное поражение сосновых лесов в ближней зоне ЧАЭС.
1 -леса; 2 - погибший "рыжий лес"; 3 - погибший "рыжий лес", раскорчеванный со
сплошным снятием грунта; 4 -луга; 5 - залежи; 6 - участки сплошного снятия грунта;
7 - застроенные территории; зоны поражения сосновых лесов: 8 летального (более 500 мР/ч);9 -сублетального (50 - 500 мР/ч); среднего (20 - 50 мР/ч)
Прирост сосны в зоне сублетального поражения, приостановившийся в 1986 г., в следующем году возобновился.
В зоне среднего поражения, которая ограничена изолинией МЭД 50 мР/ч на 01.06.86, расчетная поглощенная доза оценена в 3 - 5 Гр. Зона характеризуется угнетением роста и гибелью точек роста хвойных пород летом 1986 г. и образованием радиоморфозов, особенно сильно выразившихся в 1987 г. В этой зоне оказалось 11920 га сосновых лесов.
Оставшаяся часть 30-километровой зоны и ряд радиоактивных пятен за ее пределами могут быть отнесены к зоне слабого радиационного поражения. Внешние признаки морфологических нарушений у хвойных здесь отсутствовали, но в 1986 - 1987 гг. отмечалось снижение всхожести семян и хромосомные аномалии в мейозе.
Радиационное поражение лесных биоценозов в зоне Чернобыльской катастрофы не приобрело массового характера и коснулось преимущественно сосновых лесов. Гибель сосновых насаждений охватила менее 0,5 % площади лесов зоны, не оказала существенного влияния на радиоэкологическую обстановку в целом, хотя и ухудшила ее в непосредственной близости от атомной станции.
Гибель животных. Радиочувствительность позвоночных животных выше, чем радиочувствительность хвойных растений. Однако документальных свидетельств о массовой гибели позвоночных в ближней зоне в острый послеаварийный период нет - первые биологические наблюдения в зоне отчуждения были начаты лишь в июне-июле 1986 г.
По данным количественных учетов мышевидных грызунов в сентябре 1986 г., в ближней зоне численность этих животных снизилась в 3 - 5 раз (3,5 % ловушко-суток вместо 15 - 20 в контрольных, слабо загрязненных участках). Расчетная поглощенная доза в первый месяц после аварии составляла на учетных площадках 22 и 860 Гр для гамма- и бета-облучения соответственно [112].
Беспозвоночные на один-два порядка более устойчивы к действию ионизирующей радиации по сравнению с позвоночными. Тем не менее три экологические группы беспозвоночных сильно пострадали из-за специфических особенностей облучения в их средах обитания. Скопление радионуклидов в подстилке хвойного леса буквально выжгло мелких беспозвоночных (панцирные клещи, ногохвостки) на учетных площадках в 3 км к югу от аварийного энергоблока - их численность в июле 1986 г. упала на 2 порядка. Резко снизилась также численность почвенных беспозвоночных, лучше экранированных от бета-излучения. Соотношение неполовозрелых и половозрелых особей изменилось в пользу последних. Величина поглощенной дозы 30 Гр не влияла непосредственно на взрослых животных, но заметно сказалась на ювенильных стадиях [64]. В загрязненных лишайниках-эпифитах Hypogymnia physodes (L.) Nyl. на тех же учетных участках в 3 км к югу от блока вообще не удалось обнаружить панцирных и гамазовых клещей, ногохвосток, обычных для этой экологической ниши [75].
Изменение микробного ценоза почвы. В 1986 - 1992 гг. проводился мониторинг таксономического состава и численности почвенных микромицетов в зоне отчуждения и в Вышгородском р-не Киевской обл. Из 670 образцов на дерново - подзолистых и супесчаных почвах выделено свыше 3000 культур грибов, отнесенных после идентификации к 180 видам 72 родов.
В течение 1986 - 1988 гг. при общем снижении (в 2 - 3 раза) количества грибных диаспор и длины грибного мицелия в почве всех 7 пунктов наблюдения 30-километровой зоны отмечали преобладание темноокрашенного грибного мицелия на глубине 0-10 СЃРј. Аналогичную ситуацию констатировали также при построении корреляционных плеяд по методу П.В.Терентьева [111], отражающих сформировавшиеся в этом регионе грибные комплексы. Выявлены сложно организованные корреляционные плеяды - показатель стабильности грибных комплексов с преобладанием в их структуре меланинсодержащих видов и родов (рис.1.4.8). Обнаруженное явление квалифицируется как промышленный меланизм почвенных микромицетов [44].
В 1989 г. и позднее количество грибного мицелия приближалось к значениям, известным для "чистых" регионов [25]. Большей частью доминирующее положение по количеству грибной биомассы занимали светлоокрашенные микромицеты, что характерно для микобиоты этого региона до аварии. Однако в таксономической структуре грибных комплексов в 1989 - 1992 гг. доминирующее положение по-прежнему принадлежало темноокрашенным грибам. Эта группа грибов может быть биоиндикатором неблагополучной экологической ситуации, хотя непосредственная причина наблюдаемых изменений и прямая их связь с облучением не установлены.
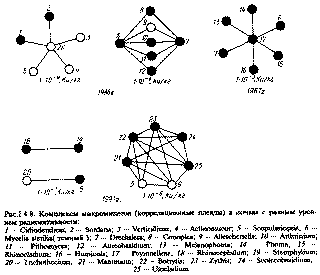
Рис.1.4.8. Комплексы микромицетов (корреляционные плеяды) в почвах с разным уровнем радиоактивности:
I -Cidiodendron; 2 -Sordaria; 3 -Verticillium; 4 -Actmomucor; 5 -Scopulanopsis; 0 - Mycelia sterilia(темный); 7 -Drechslera; 8 -Conoplea; 9 -AUescheriella; 10 -Arthrimum; II -Pithomyces; 12 -Aureobasidium; 13 -Melanophoma; 14 -Phoma; 15 -Rhinocladium- 16 -Humicola; 17 -PeyroneUeae; 18 -Rhinoccphalum; 19 -Stemphylium; 20 -Trichothecium; 21 -Mammaria; 22 -Botrytis; 23 -Zythia; 24 -Scolecobasidium; 25 -Ulocladium
4.5.3. Вторичные явления в пораженных биоценозах
Разрушение трофической пирамиды. В первые 1-3 года после гибели сосны разрушается вся система трофических связей в лесном биоценозе (рис. 1.4.9). В первую очередь лишаются пищи разнообразные насекомые, питавшиеся на хвое и побегах: личинки пилильщиков, гусеницы бабочек, тли-хермесы, сосновые клопики. С их исчезновением была подорвана кормовая база насекомоядных птиц и муравьев, собиравших в кронах белковый корм и сладкие выделения тлей. Численность почвенных микроартропод в лесной подстилке восстановилась со спадом радиационного фона [64], однако их видовой состав остался обедненным, в том числе исчезли клещи-орибатиды, промежуточные хозяева цестод, заражающих мышевидных грызунов [105].
В зоне среднего поражения в 1986 - 1987 гг. нарушалась морфология хвоинок, на 10 - 20 сут задерживался весенний рост побегов, в 2-3 раза снизился выход семян сосны, а пустозерность достигла 70 % при норме 5 - 18 % [61]. Это обедняло кормовую базу фитофагов и нарушало согласование во времени циклов развития растений и фитофагов.
Размножение насекомых-вредителей, В зоне сублетального и среднего поражения сосны возникла и сохраняется в течение последних лет вспышка массового размножения вторичных стволовых вредителей.
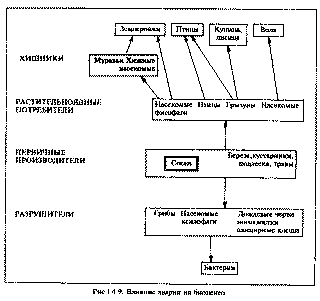
Рис.1.4.9. Влияние аварии на биоценоз
Отмечено 11 массовых видов, из них главные - большой и малый сосновый лубоеды (Blastophagus piniperda L., В.minor Hart.), синяя сосновая златка (Phaenops cyanea F.). Плотность заселения лубоедами в Лелевском и Новошепеличском лесничествах достигла 1,6 - 1,9 особей/дм2. Для дополнительного питания лубоеды внедряются в побеги. Хотя абсолютная численность вредителей возросла лишь вдвое по сравнению с нормой, но уменьшилось число побегов и резко увеличился процент заселения [I]. Опад поврежденных побегов возрос до 70 - 75 %, против 25 - 30 % в контроле. В 1990 г. практически все побеги в зоне сублетального поражения были уничтожены. Деятельность вредителей ускоряет гибель пострадавших деревьев.
Через год после гибели хвойные деревья заселялись в массовом количестве обитающими под корой жуками-трухляками Pytho depressus L. и усачами рода Rhagium. Пик их численности (несколько сотен особей на дерево) пришел на 1989 г., а уже в 1991 г. очередные погибшие деревья были слабо заражены. По-видимому, медленно засыхавшие деревья, погибшие в более поздние сроки, были мало привлекательны для стволовых вредителей из-за низкого содержания крахмала в лубе и заболони [68]. Под действием грибов и насекомых деревья полностью теряют кору и через 3 - 4 года падают.
Захламленность окружающих лесов сухостоем поддерживает кормовую базу вредителей и создает реальную опасность массового размножения их в зоне среднего поражения.
Сукцессии растительности в пораженных лесах. В зоне летального поражения произошло полное уничтожение лесных ценозов, поскольку сосна погибла, а лесные компоненты травяного покрова были подавлены вторжением видов-эксплерентов. В зонах сублетального и среднего поражения лесные ценозы сохранились. Во всех этих случаях в ценозах начались восстановительные сукцессии, отличающиеся по степени продвинутсти [12]. Гибель древесного яруса и осыпание хвои вызвали осветление леса, а также повышение уровня грунтовых вод, увеличение влажности почвы. На участках, где произошла массовая гибель сосны, по аналогии с зарастанием лесосек и гарей, это привело на первом этапе к появлению сорно-опушечных видов, таких, как ослинник двулетний (Oenothera biennis L.), мелколепестник канадский (Erigeron canadensis L.), тысячелистник (Achillea millefolium L.), марьянник обыкновенный (Melampymm vulgatum Pers.) и др.
При этом сохранились виды, доминировавшие в определенных типах сосновых боров. Например, в зеленомошниках это черника •(Vacciniwn myrtillis L.), вереск (Calluna vulgaris (L.) Hull), овсяница овечья (Festuca ovina L.), орляк (Pteridium aquilinum (L.) Kuhn), в сосняках лишайниковых - тимьян (Thimus serpillum L.), овсяница овечья, полевица тонкая (Agrostis tenuis Sibth.), ястребинка волосистая (Hieracium pfflosella L.). Также фрагментарно в виде пятен разного размера сохраняется прежний моховой и лишайниковый покров.
В дальнейшем основу травяного покрова образуют, вплоть до сплошных зарослей, типичные растения-псаммофиты: булавоносец седоватый (Corynephonis canescens (L.) Beauv.), иван-чай узколистный (Chamaertion angustifolium (L.) Holub), щавель воробьиный (Rumex acetosella L.), цмин песчаный (Helichrysum arenamim (L.) Moench), но чаще всего и в большом количестве - вейник наземный (Calamagrostis epigeios (L.) Roth).
Часто в разреженном лесу при наличии зачатков образуются также заросли малины (Rubus idaeus L.) и ежевики (R. caesius L., R-nessensis W.Hall). В образовавшихся "окнах" на месте выпавших деревьев появляются проростки деревьев. В массовом количестве в первую очередь здесь появляется молодняк березы поникшей (Betula pendula Roth.), значительно реже - осины (Populus tremula L.). Восстановлению березы способствует активная роющая деятельность кабанов (Sus scrofa L.). Количество проростков сосны по сравнению с березой в тысячи раз меньше и в первые годы жизни их прирост очень мал. Поэтому при восстановлении сомкнутого древостоя, как правило, образуется березовый (вторичный) лес, при сохранившихся соснах - сосново-березовый. Сосновый бор образуется только после отмирания березового древостоя, т.е. через много десятков или даже сотен лет.
Миграция животных в зону поражения, С соседних участков на территорию "рыжего леса" постоянно проникали зайцы-русаки (Lepus europaeus Pall.), лисицы (Vulpes vulpes L.), бродячие собаки, залетные птицы, вероятно, мигрировали и мышевидные грызуны. В марте 1987 г. во время работ по захоронению следы грызунов были обнаружены в наиболее загрязненном участке леса, у забора ЧАЭС [119]. В том же 1987 г. численность мышевидных грызунов на ст. Янов и на учетных площадках в "рыжем лесу" восстановилась за счет миграции [112].
Участки погибшего леса и пустоши над захороненным лесом постоянно посещаются крупными копытными (кабан, лось Alces alces (L.), в меньшем количестве - косуля Capreolus capreolus (L.)), однако места их отстоя, лежек или основных кормежек находятся на расстоянии 1-2 км, в зеленом хвойном лесу, вкраплениях лиственного леса и на заболоченных участках. Об этих визитах свидетельствуют следы животных.
Учет численности птиц по голосам, проведенный в мае 1987 г. в правобережном участке "рыжего леса" (мощность экспозиционной дозы составляла 40 мР/ч), показал, что прилетевшие с зимовки птицы поселились в кронах и среди мертвых деревьев в количестве до 6 семей на гектар (были зарегистрированы вертишейка (Jynx torquffla L.), серая мухоловка (Muscicapa striata (Pall.)), зяблик (Fringilla coelebs L.), полевой конек (Anthus campestris L.), белая трясогузка Motacilla alba L.)). Однако эта численность была в 4 - 5 раз ниже нормальной на контрольных участках здорового соснового бора или зеленых сосен, задержанных в росте из-за лучевого поражения [12].
Сукцессии растительности на площадях дезактивации. Из площадей, подвергавшихся дезактивации, наиболее сильно изменены участок вырубленного и захороненного леса, участок между ЧАЭС и Припят-ским затоном, где была проведена отсыпка песка, и прирусловая дамба, намытая вдоль левого берега р.Припять, захоронение дачной застройки у ст. Янов. В зависимости от степени влияния разнообразных факторов на отдельные участки дезактивированных территорий на них можно наблюдать разные стадии восстановительных сукцессии растительного покрова. Ход сукцессии рассматривается на примере "рыжего леса".
В урочище "рыжий лес" высокие деревья были срублены, а низкие захоронены в траншеи, освобожденная территория засыпана слоем песка до 0,5 м и более. После этого песчаный покров был обработан для закрепления химическими реактивами, образовавшими пленку. Поскольку она оказалась неустойчивой и препятствовала естественному зарастанию, был проведен посев ржи с подсевом смеси многолетних трав. Из видов, использованных для залужения, к 1992 г. сохранилась в заметном количестве лишь ежа сборная (Dactylis glomerataL.).
Date: 2015-09-17; view: 637; Нарушение авторских прав