Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Экономический и экологический ущерб 15 page
|
|
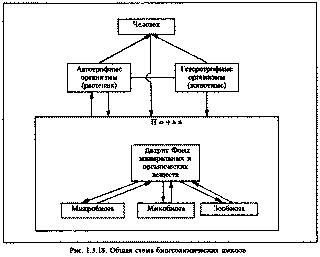
Рис. 1.3.18. Общая схема биогеохимических циклов.
Трофическая, или пищевая цепь описывает взаимодействия организмов, через которые в экосистеме происходит преобразование веществ и энергии. В описании этих взаимодействий основным является связь "пища - потребитель". В состав пищи каждого вида обычно входит не один, а несколько видов. В свою очередь каждый вид может служить пищей для многих видов. Поэтому трофические цепи образуют трофическую сеть. Поскольку экологическая система в отсутствие ее нарушений пребывает в определенном равновесном состоянии, потоки веществ характеризуются известным постоянством скоростей, которые, впрочем, подвергаются определенным сезонным колебаниям. Попавшие в экосистемы радионуклиды заполняют эти трофические сети.
Биогеохимический круговорот радионуклидов, осуществляющийся посредством функционирования трофических цепей, включает в себя:
поглощение растениями, животными, микроорганизмами отдельных радиоактивных изотопов, при этом происходит постепенное перемешивание радионуклидов с их изотопными и неизотопными носителями и их включение в состав биологических структур;
выделение надземными частями и корневыми системами растений радионуклидов в составе определенных соединений, вымывание из листьев дождями подвижных радионуклидов, например, цезия;
выделение животными продуктов, образующихся в результате пищеварения, которые поступают в почву в составе новых соединений или как их примеси;
отмирание надземных и подземных органов растений - листового опада или растений, завершивших свой онтогенез;
разложение органических остатков микроорганизмами, сопровождающееся включением радионуклидов в состав бактериальной массы или их переходом в почвенный раствор.
Различают два основных типа трофических цепей - пастбищные и детритные. В пастбищной трофической цепи основу составляют растения, которые поедаются растительноядными животными, затем следуют хищники (консументы) 1-го порядка и далее - хищники 2-го порядка. Обычно число звеньев в трофической цепи не превышает 4-5. Более длинные цепи характерны для океанических трофических цепей [5].
Вхождение радионуклидов в биомассу разных групп организмов происходит соответственно их положению в трофической цепи. Так, в водной экосистеме, которая содержит пастбищные трофические цепи, первоначально радионуклиды попадали в планктонные организмы, затем - в рыбы, питающиеся планктоном (консументы 1-го порядка), а затем - в хищники, питающиеся другими рыбами (консументы 2-го порядка). После аварии легко прослеживалась такая динамика заполнения радионуклидами трофических цепей: сначала радионуклиды появились в илах, затем - в донных рыбах, а позже - в хищниках, питающихся донными рыбами.
В детритной трофической цепи большая часть растительной биомассы не потребляется растительноядными животными, а отмирает и подвергается разложению сапрофитными организмами. В результате возникает детрит - остатки разложившейся биомассы вместе с перерабатывающими ее микроорганизмами. Детрит служит пищей так называемым детритофагам, а также является источником элементов питания для повторного питания растений. Детритофаги - дождевые черви, личинки некоторых насекомых, в водоемах - донные животные служат пищей для консументов 1-го порядка. Детритная трофическая цепь характерна для лесов, т. е. для тех экосистем, в которые были инжектированы радионуклиды. С осени 1986 г. и поныне в лесах и лесопарках, которым свойственны детритные трофические цепи, наблюдается концентрирование радионуклидов в лесной подстилке. В водоемах, где также имеются детритные трофические цепи, также регистрируется нагнетание радионуклидов в илы.
Вследствие замкнутости биогеохимического круговорота радионуклиды не должны были бы выходить за пределы определенной экосистемы. Однако в природных условиях биогенная миграция веществ, в том числе радионуклидов, сочетается с физико-химическими процессами превращений и транспорта элементов, зависящими от геохимических особенностей ландшафта. Перенос радионуклидов осуществляется, например, местным стоком, фильтрацией, зависящими от формирования химизма вод, и т.д. Помимо этих геохимических факторов, перемещение вещества, включая радионуклиды, осуществляется благодаря мигрирующим животным. Например, сезонные миграции перелетных птиц, в частности уток, сопровождаются выносом из загрязненных зон ряда радионуклидов; благодаря жизнедеятельности роющих животных радионуклиды проникают в глубь почвы значительно быстрее, чем это допускают процессы переноса радионуклидов грунтовым раствором.
В результате совокупного действия биогеохимических и физико-химических процессов происходит перераспределение радионуклидов в масштабе ландшафта. За более или менее продолжительный интервал времени установится относительное равновесное состояние распределения радионуклидов, при котором будет достигнут постоянный уровень содержания радионуклидов в биомассе отдельных звеньев трофических цепей со свойственными им сезонными колебаниям. Как уже отмечалось, помимо биогеохимического превращения радионуклидов, сопряженных с трофическими цепями, благодаря ряду геохимических процессов и таких явлений, как эрозия грунтов, смыв, формирование твердого стока, пылеперенос и т.п. всегда имеет место утечка радионуклидов из биогеохимического круговорота и расширение ареала распространения радионуклидов в среде. Со временем устанавливаются постоянные значения потоков латерального и вертикального переноса радионуклидов как показатели их утечки из конкретной трофической цепи. Такой структурой миграции радионуклидов определяется характер их концентрирования в отдельных компонентах ландшафтов.
Например, приведем схему распределения радиоактивности в лесном фитоценозе и изменения профиля распределения радиоактивности с течением времени после аварии на ЧАЭС. На землях, находящихся в сельскохозяйственном пользовании, утечка радионуклидов связана с вывозом части биомассы в виде урожая - зерна, соломы, сена, корнеплодов, другой продукции растениеводства, овощеводства и садоводства. Частично радионуклиды, отчлененные в ходе хозяйственной деятельности, возвращаются в исходные культурные фитоценозы в составе навоза или компостов. Однако возвращение радионуклидов в этом случае не может быть полным.
За годы, прошедшие после аварии на ЧАЭС, в большей части экосистем равновесие в сфере трофических цепей еще не наступило. Следует отметить, что равновесие могло бы наступить быстрее, если бы радионуклиды изначально попали в среду в растворимой форме. Однако часть активности сосредоточена в составе "горячих" частиц, которые отличаются крайне слабой растворимостью, и это замедляет усвояемость радионуклидов растениями [З].
Поскольку радионуклиды выпали на поверхность почв и на растительный покров, то они оказались сосредоточенными в самом поверхностном слое, будучи поглощенными там почвенным поглощающим комплексом или находясь в составе органического вещества отмерших осенью частей загрязненных ими растений. Зона, в которой размещена основная часть поглощающих минеральные вещества корневых систем, находится глубже места сосредоточения радионуклидов, и поэтому должно было пройти определенное время, чтобы радионуклиды могли проникнуть к корнеобитаемой зоне, что обеспечивает их корневое поступление в растения. Там, где не было вспашки, еще и поныне радионуклиды не достигли корнеобитаемой зоны, и биогеохимические циклы установились лишь в отдельных частях ценоза, например, в лишайниково-моховом ярусе.
На пахотных почвах радионуклиды были "размазаны" по профилю почвы, и корневое усвоение радиоактивных веществ установилось с
Биогеохимические циклы и миграция радионуклидов в ландшафтах. Поскольку биогенная миграция радионуклидов происходит в условиях определенных экосистем в ландшафтах и сочетается с процессами геохимического характера, которые определяют перераспределение радионуклидов между ландшафтами, следует учесть особенности и самих ландшафтов, и роль ландшафтно-геохимических барьеров.
В 30-километровой зоне основными типами ландшафтов являются элювиальные, транзитные, транзитно-аккумулятивные и аккумулятивные. Они представлены надпойменными террасами, моренными водораздельными равнинами, широкими речными долинами, в которых развиты разнообразные биогеоценозы: заболоченные луга, пойменные луга, черноолыпанник, сосняк, смешанные леса. На оставленных ранее культивируемых землях развились типичные залежи. Биогеоценозы, развившиеся в этих ландшафтах, характеризуются соответствующими трофическими цепями и отличаются определенной автономностью в отношении удерживания радионуклидов. Чем большая активность вовлечена в сферу биогеохимических циклов, тем выше степень автономности ландшафта, так как радионуклиды связаны с "живым веществом", и очень малые их количества остаются в состоянии, при котором достаточно высока их способность к геохимическим абиотическим превращениям, радиальной и латеральной миграции.
В зоне Украинского Полесья часто встречаются биогеоценозы с весьма скромной растительностью, Например, это характерно для редких сосняков на бедных песках. В этом биогеоценозе преобладают чисто геохимические процессы миграции радионуклидов. Наличие биогеохимической автономности отдельных ландшафтов, когда макро-и микроэлементы питания растений удерживаются в фитомассе или детрите, позволяет говорить о наличии ландшафтного биогеохимического барьера для ряда химических элементов, в том числе и для радионуклидов. Поскольку не все радионуклиды являются органогенными, степень проницаемости биогеохимических барьеров для разных радионуклидов может оказаться неодинаковой. Так, в настоящее время отмечают повышенную миграционную способность ^Ru. В общем же пока наблюдаются перераспределение радионуклидов, обусловленное постепенным заполнением звеньев трофических цепей, и некоторые потери радионуклидов по каналу геохимической вертикальной миграции. В последние годы отмечается переход части радионуклидов из подгоризонта лесной подстилки в подподстилочный слой.
Латеральная миграция видна только лишь на очень малых расстояниях в элементах микрорельефа. Вследствие проведенных после аварии гидротехнических работ на значительной территории изменился гидрологический режим и повысился уровень грунтовых вод, что влияет на состояние отдельных экосистем, в которых происходит изменение биоценозов. Уровень залегания грунтовых вод существенно сказывается на вертикальной миграции радионуклидов в глубь почвенного профиля.
Водные экосистемы характеризуются большой динамичностью круговорота радионуклидов. Это определяется очень сильным накоплением радионуклидов водной растительностью, фито-, зоопланктоном, большой скоростью переноса растворенных форм радионуклидов в воде [28]
Биогенные свойства радионуклидов. Основная биомасса, в которую оказались вовлеченными радионуклиды после их попадания в среду, представляет собой массу растений - фитомассу. Действительно, биомасса животных по сравнению с фитомассой очень мала. Так, в травянистых сообществах на долю животной массы в зоне Полесья Украины приходится не более 1 - 2 %. Поэтому очевидно, что степень участия радионуклидов в биогеохимических циклах определяется тем, с какой эффективностью используются в жизнедеятельности растений соответствующие тем или иным радионуклидам носители. Такая эффективность носителей определенных химических элементов называется их биогенностью.
Показателем биогенности элемента является отношение содержания элемента в живом организме к кларку биосферы. Чем выше значение биогенности элемента, тем полнее вовлекается в биогеохимические циклы радионуклид, для которого этот элемент является носителем.
Для характеристики биогенности радионуклидов используют следующие величины:
коэффициент аккумуляции (КА)
Бк/кг растительного материала
КА= -----------------------------------------
Бк/кг почвы
почвенный коэффициент пропорциональности (КЛ)
Бк/кг растительного материала
КП= --------------------------------------------------------------
Плотность поверхностного загрязнения,Бк/м2
коэффициент биологического поглощения (КБП)
Бк/кг золы растительной массы
КБП= -----------------------------------------------------------
Плотность поверхностного загрязнения,Бк/м2
коэффициент дискриминации (КД)
Активность радионуклида
(-------------------------------------) в растении
Концентрация носителя
КД=--------------------------------------------------------
Активность радионуклида
(------------------------------------)в почве
Концентрация носителя
Эти единицы характеризуют скорость вовлечения радионуклидов в растительность, как в основное звено биогеохимического круговорота веществ, и их использование позволяет количественно описывать движение радионуклидов в тех или иных экосистемах и ландшафтах.
Значения КА радионуклидов зависят от многих факторов: природы радионуклида и его физико-химического состояния; вида растений и периода его онтогенеза; типа почв и содержания в почве питательных веществ и элемента-носителя; условий увлажнения, кислотности почвенного раствора; аэрации почвы. В табл. 1.3.34 приведены пределы варьирований значений этих коэффициентов для растений. Данные являются итогом обобщения результатов определений значений этих коэффициентов на многих видах растений [22].
Значения КА радионуклидов были предметом очень обстоятельных исследований, проводимых с начала испытаний ядерного оружия и в связи с крупными ядерными авариями. Конечно, эти значенияносят только ориентировочный характер, так как они были получены в условиях экспериментов, в которых, естественно, не представлялось возможным испытать исчерпывающе широкий круг видов растений и воспроизвести условия загрязнения почв радионуклидами, которые сложились в зоне влияния аварии на ЧАЭС.
Таблица 1.3 34
Коэффициенты аккумуляции (КА) радионуклидов растениями
| Радионуклид | КА | Радионуклид | КА |
| 35S | 20 -60 | 140Ba | (2 - 5)·10-2 |
| 45Са | (4 - 6)·10-2 | 141Ce144Ce | (6 - 30)·10-4 |
| 54Mn | 0,02 - 15 | 147Pm | (3 - 30)·10-5 |
| 55Fе 59Fе | (1 - 8)·10-2 | 195W | 0,13 -0,3 |
| 60Со | (4 - 50)·10-3 | 210Pb | 0,05 - 0,43 |
| 65Zn | 3,3 - 15 | 226Ra | (1 - 40)·10-3 |
| 90Sr | 0,02 - 12 | 232Th | (1 - 700)·10-3 |
| 91Y | (3 - 70)·10-3 | 238U | 1,6·10-3- 0,1 |
| 95Zr | (3 - 80)·10-3 | 237Np | n·10-2- n·10-1 |
| 103Ru106Ru | (2 - 3)·10-3 | 239Pu240Pu | n·10-8- 1 |
| 115Cd | (4,3 - 8,5)·10-2 | 24lAm | n·10-6- 0,1 |
| 137Cs | 0,02 - 1,1 | 244Cm | n·10-4- n·10-3 |
КА радионуклидов водными растениями значительно выше, чем наземными. Не только для водорослей, но и для высших растений характерно концентрирование радионуклидов в многие сотни и тысячи раз. В известной мере это объясняется тем, что в воде разбавление радионуклидов носителями меньше, чем в почвах. Весьма высоки значения коэффициентов накопления радионуклидов и для водных животных. Например, для сома в 1987 г. в водоеме-охладителе коэффициент накопления137Cs составил 1500 [21].
Биологическое разнообразие коэффициентов накопления радионуклидов растениями. Разные виды растений очень сильно различаются по способности накапливать радионуклиды в своей биомассе. В этой их особенности находит проявление свойство растений избирательно накапливать те или иные элементы минерального питания. Те виды, которые характеризуются способностью к накоплению повышеного количества элемента, являющегося носителем для соответствующего радионуклида, естественно, - накопители последнего. Так, растения-кальциефилы накапливают большую активность радиоактивных изотопов89Sr и90Sr, а калиефилы отличаются повышенной способностью к накоплению радионуклидов134Cs и137Cs. Поэтому заведомо следует ожидать, что растения семейства мотыльковых Faboideae как кальциефилы должны накапливать в своей биомассе больше радионуклидов стронция, чем подсолнечник Helianthus annus L. и картофель Solanum tuberosum L., которые, являясь калиефилами, должны иметь повышенную склонность к накоплению радионуклидов цезия. Иначе говоря, в способности растения накапливать радионуклид основную роль играет степень потребности для организма соответствующих элементов носителей, их органогенность. Поэтому радионуклиды, относящиеся к макроэлементам питания растений, обладают относительно большей подвижностью, чем радионуклиды тех элементов, которые не относятся к элементам минерального питания растний. Вследствие этого наиболее подвижными являются радионуклиды-аналоги макроэлементов питания растений -134Cs,137Cs,89Sr,90Sr.
Вместе с тем, поведение радионуклидов других элементов, в частности тех, которые находятся в среде в рассеянном состоянии, может и не отражать степень соответствия коэффициентов накопления радионуклида и их причастность к органогенным элементам. Поведение рассеянных элементов отличается известным своеобразием, которое еще предстоит исследовать. Кроме того, радионуклиды из редкоземельных элементов, видимо, более усвояемы растениями, чем это ранее полагали. Поэтому наблюдается накопление радионуклидов цезия в фитомассе. В пределах одного участка с равномерной плотностью радионуклидного загрязнения наиболее радиоактивными являются лишайники, грибы и мхи. В некоторых случаях на лесных участках с высоким уровнем радионуклидного загрязнения содержание137Cs достигает миллионов беккерель на килограмм. Не меньшие уровни радионуклидного содержания и в талломе лишайников, которые по этой причине могут служить очень чувствительным индикаторными видами для оценки радиоактивного загрязнения территории. Например, для лишайника кладонии мягкой Cladonia silvatica, отобранной на песчаном грунте в 30-километровой зоне в 1989 г., КА радионуклидов составил 180. В 1986 г. активность его таллома была свыше 400 000 Бк/кг.
Виды грибов различаются по способности к накоплению радионуклидов. Наиболее радиоактивными в зоне влияния аварии на ЧАЭС являются такие виды: горькушка, говорушка, свинушка тонкая, волнушка розовая, ежевик выемчатый, сыроежки. Менее других видов накапливает радионуклиды белый гриб, хотя и в нем содержится намного больше радионуклидов, чем в высших цветковых растениях. Из съедобных грибов менее радиоактивными являются лисичка настоящая, подосиновик красный, дождевик жемчужный. Наиболее высокую радиоактивность наблюдают у гриба польского.
Накопление радионуклидов в грибах зависит не только от вида, но и от условия их произрастания. Например, грибы в микропонижениях рельефа накапливают радионуклидов значительно больше, чем в микроповышениях, в случае более кислых почв радиоактивность плодовых тел грибов выше, чем при меньшей кислотности почвенного раствора, в гидроморфных условиях радиоактивность плодовых тел грибов намного больше, чем в более засушливых условиях.
Хотя грибы, лишайники и мхи отличаются высокой способностью накапливать радионуклиды, их вклад в биогеохимический круговорот не очень большой, что объясняется малым значением биомассы, продуцируемой этими организмами на единицу площади.
Повышенные количества радионуклидов накапливают некоторые лесные кустарнички - черника Vasinium myrtillus L., брусника Vasinicum vitis-ideaL. и др.
Древесные растения в первое время после аварии были преимущественно механически загрязнены аэрозольными частицами. При этом наибольшая доля радиоактивности была сосредоточена в наружной коре. Даже в Киеве регистрировалась высокая удельная радиоактивность внешней коры деревьев, которая достигала в 1987 г. значений порядка десятков тыся·беккерель на килограмм. Внутренние ткани деревьев не содержали существенной активности радионуклидов, поскольку еще не могло осуществляться их корневое поглощение. Спустя 5-6 лет после аварии вследствие естественного самоочищения наружной коры стволов, радиоактивность этой части растений несколько снизилась. Вместе с тем, стала возрастать радиоактивность древесины, внутренней коры, ассимилирующих органов, что было обусловлено преимущественно корневым поступлением радионуклидов, которые достигли зоны корнеобитания.
Основным радионуклидом корневого поступления в древесных растениях является137Cs. КА радионуклидов для культурных растений обычно ниже, чем для дикорастущих. Значение КА радионуклидов зависят не только от вида растения, но и от влияния многих факторов, в частности от условий выращивания. Поэтому для одного и того же вица растения, но в разных местах произрастения КА радионуклидов могут различаться в несколько раз. Особенно сильное влияние оказывает кислотность почвенного раствора, что видно из данных следующей табл. 1.3.35.
Таблица 1.3.35
Почвенные коэффициенты пропорциональности для от кислотности почвы [19]137Cs в зависимости
| Культура | pH почвенного раствора | ||
| 4,5 - 5,5 | 5,6 - 6,5 | 6,6 - 7,5 | |
| Люцерна Medicago saliva | 0,90 | 0,20 | 0,10 |
| Клевер Trifolium arvensis L. | 0,80 - 2,90 | 0,30 | 0,10 |
| Вика Visia sativa | 1,10 - 4,50 | 0,40 | 0,20 |
| Люпин Medicago sp. | 0,90 - 2,70 | 0,30 | 0:10 |
| Горох Pisum sativum L. | 0,50 | 0,30 | 0,20 |
| Кукуруза Zea mays L. | 0,60 | 0:30 | 0,10 |
| Озимая пшеница (зерно) Triticum acstivum L. | 0,50 | 0,20 | 0:05 |
| Озимая рожь (зерно) Secalc cereale L. | 0,40 | 0,10 | 0,07 |
| Озимый ячмень (зерно) Hordcum vulgare L. | 0,30 | 0,10 | 0,00 |
| Картофель Solanum tuberosum L. | 0,30 | 0,10 | 0,04 |
| Сахарная свекла Beta vulgaris | 0,60 | 0,30 | 0,06 |
| Тыква Cucurbita pcpo L. | 0,30 | 0,10 | 0,04 |
| Огурец Cucumis | 0,10 | 0,06 | 0,03 |
| Томаты Lycopersicon | 0,20 | 0,09 | 0,03 |
| Лук AUium сера L, | 0,60 | 0,20 | 0,11 |
| Зеленые овощи | 0,30 | 0,12 | 0,02 |
| Морковь Daucus sativus | 0,30 | 0,12 | 0,05 |
Поскольку основная зона, подвергшаяся загрязнению радионуклидами, в основном содержит кислые почвы легкого механического состава с низким содержанием органического вещества и глинистых минералов, для биогеохимических процессов характерна высокая мобильность радионуклидов.
Date: 2015-09-17; view: 505; Нарушение авторских прав