Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Группы сцепления
|
|
Гены, находящиеся в одной хромосоме и наследующиеся сцепленно, составляют группу сцепления. Количество групп сцепления каждого вида должно соответствовать числу пар хромосом. Кроссинговер возникает со вполне определенной частотой для каждой пары генов, расположенных в одной группе сцепления. Причем, чем ближе в хромосоме расположены гены друг у другу, тем она выше. На основании анализа частоты кроссинговера между генами можно вычислить расстояние между генами и, таким образом, определить их локализацию в хромосоме План расположения генов в хромосоме называется картой хромосомы.
Хромосомные карты построены для животных ряда видов, однако для собаки их нет, так как для их составления необходимы фундаментальные исследования генетики вида и большой статистический материал, получение которого у собак затруднительно. Для кошки установлено положение на хромосомах приблизительно для 50 генов (Бородин, 1995).
На основании анализа результатов многочисленных экспериментов с мухой дрозофилой Т. Морган сформулировал свою хромосомную теорию наследственности, сущность которой заключается в следующем:
1. Материальные носители наследственности — гены — находятся в хромосомах, располагаются в них линейно на определенном,расстояниидруготдруга.
2.Гены, расположенные в одной хромосоме, наследуются сцепленно. Число групп сцепления соответствует гаплоидному числу хромосом.
3.Признаки, гены которых находятся в одной хромосоме, наследуются сцепленно.
4. В потомстве гетерозиготных родителей новое сочетание генов, расположенных в одной паре хромосом, может возникнуть в результате кроссинговера в процессе мейоза.
5. Частота кроссинговера, определяемая по проценту кроссоверных особей, зависит от расстояния между генами.
6. На основании линейного расположения генов в хромосоме и частоты кроссинговера как показателя расстояния между генами можно построить карты хромосом.
Сцепление генов приводит к одновременному наследованию блока признаков. Эти блоки могут сохраняться на протяжении нескольких поколений. Именно они способствуют передаче сложных признаков, например определяют внешнее сходство дедов и внуков, или обусловливают то, что все метисы колли или кокеров имеют совершенно определенную внешность независимо от внешнего вида второго родителя.
Сцепление признаков может происходить в силу целого ряда причин, среди них и сцепление генов и плейотропия и действие определенных каналов морфогенеза. Сцепление признаков в значительной степени обусловливают тип собаки, препотентность производителя и делают возможным существование заводских линий.
Кроссинговер. Генетическое доказательство перекреста хромосом. Генетические карты хромосом и принципиальный метод их получения. Схематическое изображение явления кроссинговера. Понятие о цитологических и рестрикционных картах.
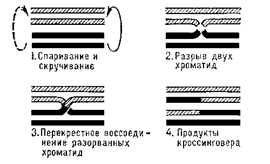
Кроссинговер — перекрест, взаимный обмен гомологичными участками гомологичных хромосом в результате разрыва и соединения в новом порядке их нитей — хроматид; приводит к новым комбинациям аллелей разных генов. Важнейший механизм, обеспечивающий комбинативную изменчивость в популяциях и тем самым дающий материал для естественного отбора. Протекает в мейотически, реже — в митотически делящихся клетках. Может приводить к перекомбинации больших участков хромосомы с несколькими генами или частей одного гена (внутригенный кроссинговер), обеих нитей молекулы ДНК или только одной. Частота кроссинговера между генами отражает расстояние между ними в хромосоме. Иными словами, в паре гомологичных хромосом между несестринскими хроматидами происходит обмен гомологичными участками. Поскольку в паре хромосом одна хромосома происходит от матери, а другая — от отца, процесс кроссинговера ведет к внутрихромосомным рекомбинациям наследственности. Молекулярный механизм кроссинговера окончательно не выяснен.
Возможность влияния на процесс кроссинговера. Хромосомы являются неотъемлемой частью всей системы клетки. Редупликация хромосом и весь цикл деления являются результатом деятельности всей клетки.
Перекрест хромосом как сложный физиологический, физический и биохимический процесс подвержен влиянию факторов внешней среды, функционального состояния целого организма и отдельной клетки. Перекрест хромосом обусловлен также их внутренней структурой и генным составом, т. е. генотипом.
Перекрест у гомои гетерогаметного пола. У большинства изученных животных и растений хиазмы и перекрест осуществляются в мейозе у обоих полов. Но существуют отдельные виды животных, у которых мейотический кроссинговер происходит только у гомогаметного пола, а у гетерогаметного в норме отсутствует. Это касается не только половых хромосом, отсутствие синапсиса у которых можно было бы объяснить их гетероморфностью у гетерогаметного пола, но и аутосом.
По некоторым цитологическим данным, у самцов дрозофилы и самок шелкопряда в профазе I мейоза либо полностью отсутствует синапсис и образование хиазм, либо эти процессы происходят не в профазе мейоза, а-на более ранних стадиях развития половых клеток (на стадии сперматогониев и оогониев). У гомогаметного пола этих видов (самок дрозофилы и самцов шелкопряда) перекрест хромосом протекает нормально.
У многих видов млекопитающих, птиц, рыб и насекомых гетерогаметность пола не сказывается на процессе кроссинговера.
Влияние структуры хромосом на частоту перекреста. Известно, что хромосома состоит из эухроматиновых и гетерохроматиновых участков. Целостная структура хромосомы в митозе и мейозе обеспечивается наличием центромеры. Около центромеры имеются, как правило, гетерохроматиновые участки.
Центромера играет очень важную роль и в механизме перекреста хромосом.
У дрозофилы в норме вблизи центромеры перекрест происходит редко. По мере удаления от нее частота перекреста увеличивается, а затем к концам снова уменьшается. Поэтому на генетических картах дрозофилы вблизи центромеры локализуется генов больше, чем в удаленных от нее районах. У кукурузы такого влияния центромеры на кроссинговер не обнаружено.
На частоту перекреста в разных участках хромосомы влияет и распределение гетерохроматиновых и эухроматиновых районов. Гетерохроматиновые участки способствуют высокой изменчивости частоты перекреста под влиянием факторов внешней среды. Значение гетерохроматиновых и эухроматиновых районов для кроссинговера может быть связано со степенью спирализации хромосомной нити в этих районах. Понижение частоты кроссинговера в центромерном районе может быть связано с высокой спирализацией этого района хромосомы. Спирализация уменьшает цитологическое расстояние между генами, а ее усиление может препятствовать синапсису гомологичных районов и перекресту. 164
Влияние функционального состояния и генотипа организма на перекрест хромосом. Частота кроссинговера зависит от возраста организма (это установлено для дрозофилы). Если изучить перекрест между двумя генами, локализованными на генетической карте на небольшом расстоянии, скажем на расстоянии 6% Друг от друга, и учитывать процент кроссинговера у самок дрозофилы по десятидневкам откладки оплодотворенных яиц (за 1 —10, 11—20, 21—30 дней), то частота перекреста будет составлять соответственно 5,9; 1,8 и 3,8%. Следовательно, первый возраст соответствует максимуму, второй — спаду, а третий — подъему процента перекреста.
Вполне возможно влияние функционального состояния организма на течение различных стадий мейоза (а значит, и на перекрест хромосом), ибо степень спирализации хромосом, скорость прохождения различных стадий профазы может в сильной степени зависеть от физиологического состояния клеток. Это тем более вероятно, что в зависимости от физиологического состояния клеток могут изменяться соотношения химических ингредиентов хромосом и цитоплазмы, а также дисперсность коллоидов, повышаться или понижаться вязкость хромосом и окружающей плазмы.
Генотип может влиять на частоту кроссинговера различными путями. Например, на частоту перекреста, в сильной степени влияют хромосомные перестройки, поскольку они нарушают нормальный синапсис хромосом в стадии зигонемы.
У кукурузы открыты гены, которые контролируют синапсис хромосом в профазе I мейоза (асинаптический ген), спирализацию и слипание хромосом; найден ген, препятствующий редукции хромосом в мейозе. У ржи также обнаружен генетический контроль частоты образования хиазм. Очевидно, что подобного типа гены могут влиять и на частоту обменов в гомологичных хромосомах.
Влияние факторов внешней среды на перекрест. До сих пор
мы рассматривали зависимость кроссинговера от генотипа и определяемого им физиологического состояния клеток и организма. Такой тип перекреста называют спонтанным кроссинго- вером. Но частоту перекреста можно изменить влиянием на организм различных факторов внешней среды.
Перекрест, вызванный искусственно влиянием различных факторов, называют индуцированным кроссинговером.
На перекрест хромосом влияют многие факторы внешней среды: высокая и низкая температура, ионизирующие излучения, присутствие в клетках ионов кальция и магния и др.
Например, у дрозофилы низкие (9—13°) и высокие (30—32°) температуры увеличивают процент кроссинговера; в оптимальных температурных условиях развития обнаруживается наименьший процент перекреста.
Дальнейшими исследованиями было установлено, что районы хромосом вблизи центромеры более отзывчивы на внешние воздействия, чем удаленные от нее. Это явление связывают с более высокой реактивностью гетерохроматиновых районов вблизи центромеры.
Ионизирующая радиация также влияет на кроссинговер, увеличивая его частоту. Уже говорилось о том, что у гетерогаметного пола (самцы дрозофилы и самки тутового шелкопряда) перекрест хромосом не обнаруживается. Однако если эти организмы подвергнуть действию рентгеновых лучей, то в потомстве возникают кроссоверные особи.
Исследование действия химических агентов также показало, что многие из них также увеличивают частоту кроссинговера. ЭДТА — этилендиаминтетрауксусная кислота—наиболее изученная из химических агентов по эффекту на кроссинговер.
Предполагается, что этот агент удаляет из хромосомы ионы кальция и магния, которые, по-видимому, играют роль в поддержании структурной целостности хромосом, поэтому удаление их ведет к нарушению непрерывности структуры хромосом, что и увеличивает частоту хроматидных разрывов, часть из которых может приводить к рекомбинации генов.
Конкретный механизм действия внешних факторов на кроссинговер все еще остается невыясненным.
Изучение сцепленного с полом наследования и явления кроссинговера дало возможность сформулировать хромосомную теорию наследственности, в первоначальном виде предложенную Морганом. Согласно этой теории единицы наследственности гены расположены в хромосомах в определенной линейной последовательности, причем каждый ген имеет в хромосоме свое определенное место —локус. Таким образом, хромосомы по своей длине наследственно дискретны. Частота кроссинговера между генами является функцией расстояния между ними.
К сказанному следует добавить, что благодаря перекресту гены могут комбинироваться и давать сочетания признаков, которые наилучшим образом обеспечивают приспособление организма к среде. В случае сцепления благоприятных генов с вредными с помощью перекреста они могут разъединяться. Таким образом, кроссинговер играет исключительно важную роль в процессе эволюции, повышая комбинативную изменчивость.
Date: 2015-09-02; view: 2147; Нарушение авторских прав