Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Оптичні властивості бактеріородопсину та особливості структурної організації пурпурних мембран
|
|
ЗМIСТ
ВСТУП…………………………………………………………………… 2
РОЗДІЛ 1. ОГЛЯД ЛIТЕРАТУРИ ПО ОПТИЧНИМ ВЛАСТИВОСТЯМ БАКТЕРIОРОДОПСИНУ…………………………………. 3
1.1. Оптичні властивості бактеріородопсину та особливості організації пурпурних мембран…………………………………………………. 3
1.2. Фотохімічний цикл реакцій бактеріородопсину…………..…….. 8
1.3. Вплив хімічних домішок на властивості плівок на основі бактеріородопсину..........................................................………………………. 9
РОЗДІЛ 2. МЕТОДИКА ОТРИМАННЯ ПЛІВОК ТА ДОСЛІДЖЕННЯ ТА ЇХ ХАРАКТЕРИСТИК………………………………….16
2.1. Методи отримання плівкових структур на базі БР……………….. 16
2.2. Схема установки для голографічного запису інформації на плівках бактеріородопсину………………………………………………………….….. 18
2.3.Механізм фотоперетворення в плівках БР під дією світла………. 20
2.4.Запис голограм в плівках БР із різними хімічним складом та залежність дифракційної ефективності від концентрації домішок…………. 22
ВИСНОВКИ ………………………….………………………………… 26
ЛІТЕРАТУРА ……………………………………..………………….… 27
ВСТУП
Сучасний стан фізики твердого тіла, напівпровідників і діелектриків характеризується суттєвою інтенсифікацією вивчення фізичних властивостей органічних матеріалів. Тенденція зменшення розмірів і великої інтеграції сучасних приладів, зростання їх економності, по новому ставить задачі вибору вихідних органічних матеріалів, придатних для створення елементів оптоелектроніки різного функціонального призначення. Завдяки своїм унікальним оптичним властивостям, необмеженим ресурсам, простоті та екологічній чистоті одержання бактеріородопсин, з моменту відкриття його у пурпурних мембранах галофільних бактерій, знаходиться у центрі досліджень, що проводяться в цьому напрямку.
Бактеріородопсин (БР) - білковий комплекс з хромофорною групою ретиналь, здійснює перетворення світлової енергії в межах одної молекули [1]. Особливості діелектричних плівкових структур на основі БР, такі як надзвичайно висока роздільна здатність, відсутність періоду схованого зображення і необхідності прояву, швидкодія, чутливість до всіх параметрів світлового випромінювання та висока циклічність надають їм перспективність практичного застосування в якості середовища для запису та обробки оптичної інформації.
Метою роботи є дослідження впливу хімічних домішок на ефективність голографічного запису інформації на плівках бактеріородопсину, зокрема дослідження залежності дифракційної ефективності від концентрації триетаноламіну.
Для виконання даної роботи необхідно виконати наступні завдання: отримати плівкові структури різного хімічного складу; дослідження дифракційної ефективності; встановити залежність дифракційної ефективності від концентрації домішок;
РОЗДІЛ 1. ОГЛЯД ЛIТЕРАТУРИ ПО ОПТИЧНИМ ВЛАСТИВОСТЯМ БАКТЕРIОРОДОПСИНУ.
Історія досліджень бактеріородопсину яскраво ілюструє швидкий, вибухоподібний розвиток сучасних напрямків фізико-хімічної біології, біоенергетики та фотобіології. Якщо до 70-х років не існувало не тільки проблеми бактеріородопсину, але й самої назви цього пігменту, то в останні роки його дослідженням займаються сотні дослідників різних спеціальностей у багатьох лабораторіях світу. Бактеріородопсин, будучи структурно близьким до зорового родопсину, має функцію не фоторецептора, а фотоенергетичного перетворювача, який забезпечує запас енергії у доступній для живої клітини формі. Відносна простота устрою цього білка дозволяє проводити дослідження фотоіндукованих процесів, що мають велику практичну цінність.
Оптичні властивості бактеріородопсину та особливості структурної організації пурпурних мембран
У життєвих явищах клітин особливо важливу роль відіграють мембрани, які обмежують клітини та її внутрішні частки. Зв’язок з навколишнім середовищем клітина здійснює через мембрану, де відбувається обмін речовин, передача інформації, перетворення енергії світла в хімічну енергію, що є життєво необхідним для всіх живих організмів. Процес перетворення накопиченої з фотосинтезу енергії живлення в АТФ відбувається за допомогою систем, функціонуючих у тісному зв’язку з мембранами [6]. Мембранні енергоперетворюючі системи поділяються на два типи:
а) системи, що перетворюють енергію світла (фотосинтетичні мембранні системи, характерні для бактерій та вищого роду рослин);
б) мембранні системи, що перетворюють енергію живлення (окислювальна фосфориляція, характерна для аеробних живих організмів).
На початку 70-х років Д. Остерхельтом і В. Стокеніусом [1] була відкрита нова енергоперетворююча система – бактеріородопсин. БР концентрується у частинах мембрани клітини Halobacterium halobium (Рис.1.1), які, завдяки їх кольору, назвали пурпурними мембранами (ПМ). Halobacterium halobium - бактерії, що живуть у дуже солоних частинах морів. Свою енергію ці одноклітинні отримують методом окисної фосфориляції в середовищі, багатому на поживу та кисень, але в випадку несприятливих умов вони спроможні використовувати енергію світла за допомогою ретиналь білкового комплексу – бактеріородопсину.
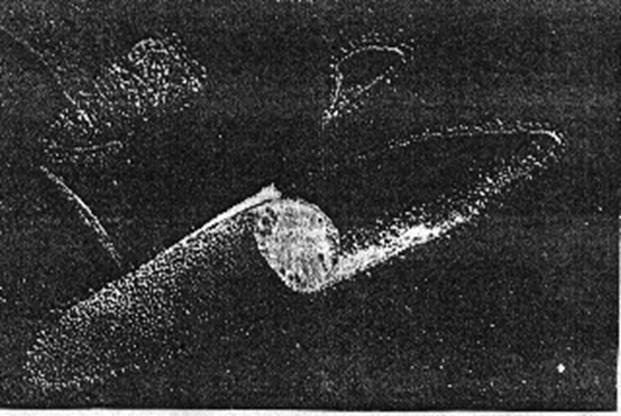
Рис. 1.1. Мікрофотографія замороженої клітини Halobacterium halobium з тріщиною (збільшення 57400) [1].
Під впливом поглинання фотону молекули БР перекачують протон із клітини в зовнішній простір, виникає різниця концентрацій протонів між двома сторонами мембрани, яка вирівнюється шляхом перекачування протонів знов у клітину через АТФ-фазу для утворення АТФ, необхідної для підтримки життєздатності клітини. Згідно хеміосмотичної теорії Мітчелла [8] енергія світла спочатку перетворюється в електрохімічну енергію протонів, а потім у енергію зв’язку пірофосфату АТФ:

 енергія світла mH+ АТФ
енергія світла mH+ АТФ
БР АТФ-фаза
Пурпурні мембрани, що складаються на 75 % з БР та на 25% з ліпіду, можна легко виділити, при цьому вони залишаються повністю стабільними, як у структурному, так і в функціональному плані [9]. Розподіл електронної густини у ПМ показаний на рис. 1.2.
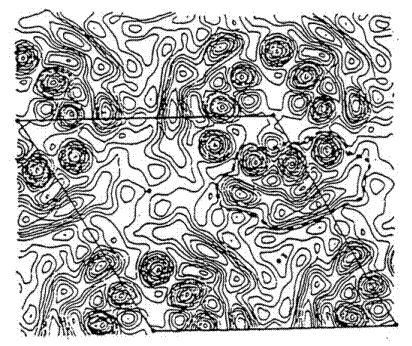
Рис. 1.2. Електронно-дифракційна картина ПМ знята зі сторони нормалі до площини мембрани [10]. Пунктирна лінія обмежує одну молекулу бактеріородопсину, суцільна лінія вказує на елементарну комірку гексагональної гратки.
Молекули БР у пурпурних мембранах, число та розмір яких залежить від умов культивування, утворюють двовимірну гексагональну гратку таким чином, що у вузлах кристалічної гратки знаходяться тріади молекул БР. Середній діаметр ПМ – 0.5 мкм, товщина – 4.9 нм [11]. Подібно до інших біологічних мембран, поверхня ПМ заряджена негативно – 0.02 зарядів / нм2, що означає 2 електрони у розрахунку на одну молекулу БР. Дипольний момент – 3.33 × 10-23 Кл×м, виникає в результаті асиметричного розподілу поверхневого заряду[12,13].
Бактеріородопсин є одним із перших мембранних білків, для якого була визначена повна амінокислотна послідовність та тривимірна структура [14, 15]. Молекула БР складається з апопротеїну, так званого опсину, та з хромофору ретиналю, його поліпептидний ланцюг складається з 248 амінокислот.
Схематична структура бактеріородопсину показує й шлях транслокації протону (рис. 1.3). Показані частини, на які ділиться молекула БР з її функціональної сторони: зовнішньо клітинна частина, де вивільнюється протон; частина, де розташована Шифова основа; та внутрішньоклітинна частина, де здійснюється захоплення протона [18-20]. Зовнішньо клітинна сторона каналу гідрофільна та набагато ширша за сторону між ретиналем та гідрофобною цитоплазматичною стороною.
Про положення ретиналю в білку можна судити за результатами досліджень кругового дихроїзму та нейтронодифракційного аналізу. Ці експерименти свідчать про те, що тяжі обмежують простір у мембранному бішарі, який містить повністю транс-ретиналь, під кутом 20о до площини мембрани, площина хромофору перпендикулярна до площини ПМ [20]. Нітроген Шифової основи розташований між C- та G- тяжами, циклогексанове кільце розташоване близько до тяжу E [17], на відстані 0.9 нм від зовнішньо клітинної сторони, Шифова основа - на відстані 1.6 нм.
Date: 2015-07-27; view: 821; Нарушение авторских прав