Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Поняття про рухову одиницю
|
|
Структурною одиницею м’язу є м’язове волокно. Функціональна одиниця м’язу отримала назву рухової (моторної) одиниці.
Рухова одиниця складається з  -мотонейрона передніх рогів спинного мозку і всіх м’язових волокон, що іннервуються цим нейроном. При збудженні мотонейрона відбувається одночасне скорочення всієї моторної одиниці.
-мотонейрона передніх рогів спинного мозку і всіх м’язових волокон, що іннервуються цим нейроном. При збудженні мотонейрона відбувається одночасне скорочення всієї моторної одиниці.
Класифікація рухових одиниць:
1 За функцією.
За цією ознакою розрізняють швидкі і повільні рухові одиниці. Для швидких рухових одиниць характерні дуже швидкі і сильні скорочення. Вони забезпечують біг на короткі дистанції (спринтери), різні види стрибків. Для повільних рухових одиниць характерні повільні, але тривалі скорочення. Вони забезпечують біг на довгі дистанції (стайєри), підтримання пози тіла. Порівняльна характеристика швидких і повільних моторних одиниць наведена у таблиці.
Порівняльна характеристика швидких і повільних рухових одиниць.

2 За будовою.
За цією ознакою розрізняють малі і великі рухові одиниці. До складу малих рухових одиниць входить невелика кількість м’язових волокон. Наприклад, у м’язах гортані рухові одиниці складаються із 2-3 м’язових волокон, у м’язах обличчя - 10-12. Як правило, малі рухові одиниці збуджуються слабкими нервовими стимулами, оскільки збудливість їх нейронів висока. До складу великих м’язових одиниць входить багато м’язових волокон. Наприклад, у м’язах кінцівок і тулуба їх кількість доходить до кількох сотень. Великі рухові одиниці збуджуються більш сильними стимулами порівняно з малими, оскільки збудливість їх нервових волокон нижча.
Фізіологічні характеристики скорочення м’язів
Навантаження
Залежно від типу навантаження можливі 2 режими скорочення ізольованого м’яза:
- ізотонічне скорочення. Цей тип скорочення виникає тоді, коли м’яз переміщує вантаж. Один кінець м’яза фіксований, інший – вільний, зв’язаний з вантажем;
- ізометричне скорочення. Цей тип скорочення виникає тоді, коли обидва кінці м’яза фіксовані.
Порівняльна характеристика режимів скорочення.

Коли йдеться не про ізольований м’яз, а про м’яз, який функціонує в організмі, то розрізняють такі види скорочень:
- ауксотонічне скорочення. Це тип скорочення, при якому одночасно зменшується довжина м’яза і зростає його напруження. Цей динамічний тип скорочення забезпечує переміщення тіла у просторі та окремих його частин стосовно одна до одної;
- ізометричне скорочення. Це тип скорочення, при якому зростає напруження м’язів без зміни їх довжини. Цей статичний тип скорочення забезпечує скорочення, спрямовані на підтримку пози тіла.
Сила
Сила, яку розвиває м’яз при скороченні, дорівнює сумі сил окремих м’язових волокон.
Розрізняють максимальну і абсолютну силу скорочення.
Максимальна сила в ізотонічному режимі визначається максимальною масою вантажу, який піднімає м’яз при скороченні; в ізометричному режимі - напруженням, яке розвиває м’яз. Максимальна сила виражається у кг. Максимальна сила залежить від будови м’яза, його функціонального стану. Наприклад, собака м’язами щелепи може підняти вантаж, у 8,3 раза більший від своєї маси. Проте максимальна сила не дозволяє порівняти силу різних м’язів. Для цього служить інший показник.
Абсолютна сила скорочення – це відношення максимальної сили ізометричного скорочення до площі фізіологічного поперечного перерізу м’яза. Абсолютна сила виражається у кг/см2. Фізіологічний поперечний переріз м’яза є площею поперечного перерізу волокон, які складають цей м’яз. Як відомо, розрізняють паралельно-волокнисті, веретеноподібні, перисті м’язи.

Типи будови різних м’язів: А – м’язи з паралельним розміщенням волокон; В – веретеноподібні м’язи; С – перисті м’язи.
Фізіологічний поперечний переріз збігається з геометричним лише у паралельно-волокнистих м’язах. У веретеноподібних і, особливо, у перистих він значно більший за геометричний.
М’язи з косим розміщенням волокон мають більший фізіологічний поперечний переріз, а отже більшу силу.
Абсолютна сила деяких м’язів наведена у таблиці.

Чинники, які визначають силу м’язового скорочення:
І На молекулярному рівні сила скорочення визначається кількістю утворених комплексів між голівками міозину і активними центрами актинових філаментів.
У свою чергу, кількість утворених актоміозинових комплексів залежить від:
- Вихідної довжини саркомера.
Максимальну силу скорочення розвиває м’яз при довжині саркомера 2,0 - 2,25 мкм. Саме при такій довжині кількість актоміозинових комплексів найбільша (рис. 9.11 А).
Якщо довжина саркомера менша за 2,0 мкм, то актинові філаменти заходять одна за одну і заважають утворенню актоміозинових комплексів.
Якщо довжина саркомера більша за 2,25 мкм, зона перекриття актинових і міозинових філаментів зменшується, а отже, зменшується і кількість голівок міозину, яка може прикріпитися до активних центрів актину (рис. Б,В).

Співвідношення між довжиною саркомера і кількістю актоміозинових комплексів.
- Кількості відкритих активних центрів актинових філаментів, яка визначається концентрацією іонів Са2+ у саркоплазмі.
Чим більше іонів Са2+, тим більше утворюється комплексів між тропоніном С і Са2+, більше відкривається активних актинових центрів, більше утворюється актоміозинових комплексів і більша сила скорочення.
- Можливості голівок циклічно від’єднуватися від одного активного центру і приєднуватися до іншого. Ця можливість залежить від наявності АТФ.
Таким чином, основними факторами, що визначають силу скорочення на молекулярному рівні, є вихідна довжина саркомера, концентрація іонів Са2+ у саркоплазмі, наявність АТФ.
ІІ На клітинному рівні сила скорочень залежить від кількості міофібрил у м’язовому волокні.
Чим більше міофібрил, тим більша сила скорочення. При посиленій роботі м’яза виникає його гіпертрофія, яка характеризується зростанням кількості міофібрил і збільшенням сили скорочення. Механізм виникнення гіпертрофії пояснює теорія Ф.Меєрсона, згідно з якою, при посиленій роботі м’яз споживає більше АТФ, у результаті гідролізу якої утворюється АДФ. АДФ є регулятором процесу транскрипції в ядрі. Внаслідок посилення процесів транскрипції і трансляції збільшується біосинтез структурних білків, зростає кількість міофібрил.
ІІІ На органному рівні зростання сили скорочення зумовлене явищами просторової і частотної сумації.
Просторова сумація – це збільшення сили скорочення м’яза за рахунок втягування у цей процес більшої кількості рухових одиниць. При слабких стимулах скорочуються лише малі рухові одиниці м’яза. При збільшенні інтенсивності стимулу у процес скорочення починають втягуватися більші рухові одиниці. Чим більше рухових одиниць збуджується, тим більша сила скорочення всього м’яза.
Частотна сумація (тетанізація) - це збільшення сили скорочення м’яза за рахунок зростання частоти стимуляції. При збільшенні частоти стимуляції до 50 імпульсів за секунду сила м’язового скорочення зростає у 3 – 4 рази.
3 Тривалість
За тривалістю скорочення бувають поодинокі і тетанічні.
Поодинокі скорочення виникають за умови, коли на одне м’язове волокно впливають одним стимулом.
У поодинокому м’язовому скороченні виділяють три фази
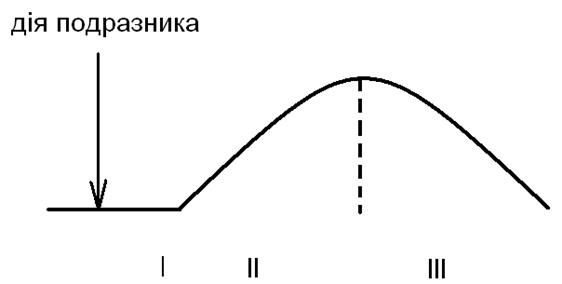
Крива поодинокого скорочення: І – латентний період; ІІ – період скорочення; ІІІ – період розслаблення.
І фаза – латентний період (час від початку дії подразника до початку скорочення м’яза), який триває у середньому 15 мс;
ІІ фаза – період скорочення (поступове зростання напруження м’яза), який триває у середньому 50 мс;
ІІІ фаза – період розслаблення (поступове зменшення напруження м’яза), який триває у середньому 50 мс.
Тетанічні скорочення (тетанус) – тривалі і сильні скорочення м’язів, пов’язані зі збільшенням частоти їх стимуляції. Для виникнення тетанусу кожний наступний стимул має діяти на м’яз, який не закінчив цикл скорочення-розслаблення.
Основу тетанусу складає явище сумації (накладання одного скорочення на інше). Щоб відбулася сумація, треба аби проміжок між стимулами був не менший за рефрактерний період м’язового волокна.
Залежно від частоти стимуляції розрізняють 2 види тетанусу: 1) з убчастий, виникає при низькій частоті стимуляції (5–50 Гц), коли кожен наступний стимул припадає на фазу розслаблення попереднього скорочення;
2) гладкий (суцільний), виникає при високій частоті стимуляції (більше 50 Гц), коли кожен наступний стимул припадає на фазу скорочення попереднього скорочення.

Види тетанусу.
Під час тетанусу зростає сила м’язового скорочення. Причиною цього є зростання концентрації іонів Са2+ у саркоплазмі, оскільки під час розслаблення він не встигає відкачуватись у СПР.
Слід пам’ятати, що під час тетанусу складаються скоротливі відповіді, але не ПД.

Одночасна реєстрація амплітуди скорочень і ПД.
4 Швидкість
Швидкість скорочення м’яза визначається двома чинниками:
- типом моторної одиниці (швидкі чи повільні).
Для швидких рухових одиниць характерні дуже швидкі і сильні скорочення, а для повільних - повільні, але тривалі. Це пояснюється тим, що максимальна швидкість укорочення саркомера (без навантаження) дорівнює максимальній швидкості ковзання актинових і міозинових ниток одна відносно одної. Що швидше поперечні містки розщеплюють АТФ і взаємодіють з актином, то більша швидкість ковзання. У повільних моторних одиницях міозин має низьку АТФ-азну активність, тому описаний процес відбувається у них повільно;
- величиною навантаження на нього.
Швидкість скорочення обернено пропорційна навантаженню на м’яз. Графічно ця закономірність має назву кривої “навантаження-швидкість” або “сила-швидкість” (крива Хіла).

Крива Хіла.
Робота
Робота м’яза вимірюється добутком маси піднятого вантажу на величину укорочення м’яза.
Розрізняють 2 види роботи м’язів:
- зовнішню (динамічну);
- внутрішню (статичну).
Зовнішня робота – це робота щодо переміщення вантажу, тіла або окремих його частин у просторі. Вона здійснюється в експериментальних умовах при ізотонічному режимі скорочення, в умовах організму при ауксотонічних скороченнях.
Зовнішня робота залежить від таких чинників:
1) швидкість скорочення. Максимальна зовнішня робота виконується при середній швидкості (закон середніх швидкостей);
2) навантаження. Максимальна зовнішня робота виконується при середніх навантаженнях (закон середніх навантажень).
Статична робота – це робота, при якій м’язові волокна розвивають напруження, але не змінюють своєї довжини. Вона здійснюється в ізометричному режимі (наприклад, робота з утримання вантажу). Внутрішня робота пов’язана з процесами, що розвиваються у самому м’язовому волокні (механічна робота щодо подолання тертя, осмотична робота щодо переміщення іонів і т. ін.).
Стомлення
Стомлення - тимчасове зниження працездатності м’яза, що настає внаслідок роботи і зникає після відпочинку.
Стомлення виявляє себе зниженням сили (амплітуди) скорочень, зростанням латентного періоду і тривалості фази розслаблення.
Чинники, які спричиняють стомлення:
І На клітинному рівні (в умовах in vitro в ізольованих м’язах):
- зниження запасів АТФ (відбувається внаслідок зменшення запасів глікогену, порушенні ресинтезу АТФ і креатинфосфату);
- накопичення продуктів метаболізму. Кислі продукти обміну (молочна кислота, піровиноградна кислота, фосфорна кислота та ін.) дифундують у навколоклітинний простір і зменшують збудливість м’язового волокна, пригнічують гліколіз і утворення АТФ, конкурентно зв’язуються з тропоніном С, витісняючи кальцій.
Слід зазначити, що стомлення ізольованого скелетного м’яза при його прямому подразненні є лабораторним феноменом. В умовах організму до м’яза постійно надходить кров, яка постачає необхідні поживні речовини і забирає продукти обміну. Тому in vivo стомлення рухового апарату при тривалій роботі розвивається більш складно і залежить від інших чинників.
ІІ На органному рівні (в умовах in vivo):
- стомлення відповідних нервових рухових центрів;
- стомлення нервово-мязових синапсів.
Доказом того, що зниження працездатності м’язів перш за все пов’язане зі стомленням нервових центрів, є досліди І.М.Сєченова, у яких він довів, що відновлення працездатності стомлених м’язів руки людини відбувається швидше, якщо у період відпочинку працює інша рука або м’язи нижніх кінцівок. Такий відпочинок І.М.Сєченов назвав активним.
Доведено, що м’язи, які виконують статичну роботу, стомлюються швидше, ніж м’язи, які виконують динамічну роботу. Наприклад, людині важче стояти, ніж ходити.
Для вивчення стомлення м’язів у людини у лабораторних умовах користуються ергографами – приладами для запису механограми при рухах, які ритмічно виконуються групою м’язів. Найпростішим ергографом є ергограф Моссо. Прилад записує амплітуду рухів пальця руки, до якого прикріплений вантаж. Запис кривої стомлення називається ергограмою.

Ергограма, зафіксована на ергометрі Моссо.
Існують ергографи, які фіксують ті чи інші рухи людини. Так, у клініці широко використовують велоергографи (велоергометри). Пацієнт ногами рухає педалі приладу при певному заданому опору руху. Спеціальні датчики реєструють параметри руху, а також показники дихання, кровообігу, ЕКГ. Велоергометрія дозволяє визначити функціональні можливості організму людини.
ФІЗІОЛОГІЯ ГЛАДЕНЬКИХ М’ЯЗІВ
Гладенькі м’язи входять до складу внутрішніх органів і забезпечують моторну функцію цих органів (травна система, сечостатева система, кровоносна система та ін.).
Структурні особливості гладеньких м’язів
Гладенькі м’язи складаються із гладеньком’язових волокон (клітин). Довжина цих волокон становить 20 – 500 мкм, що в тисячі разів менше порівняно з волокнами скелетних м’язів, діаметр 2 – 5 мкм, що у 20 разів менше, якщо порівняти зі скелетними м’язами. Волокна з’єднуються одне з іншим за допомогою нексусів, через які електричні імпульси передаються від однієї клітини до іншої.
Гладеньком’язові клітини (ГМК) мають веретеноподібну форму, вкриті плазматичною мембраною.

Організація скоротливого апарату гладеньких м’язів.
На відміну від скелетних вони не мають поперечної посмугованості, що свідчить про іншу організацію скоротливого апарату.
Скоротливий апарат гладеньких м’язів, як і скелетних, складається із двох типів філаментів: тонких актинових і товстих міозинових.
Актинові філаменти складаються лише з одного білка – актину і на відміну від скелетних не містять тропоніну і тропоміозину. Актинові філаменти кріпляться до так званих щільних тілець, які відіграють роль Z- пластинок скелетних м’язів. Певного порядку у розміщенні щільних тілець немає: одні з них кріпляться до мембрани ГМК, інші розкидані у цитоплазмі. Щільні тільця сусідніх клітин можуть бути зв’язані міжклітинними протеїновими містками, завдяки чому скорочення передається від однієї клітини до іншої.
Міозинові філаменти розміщуються між пучками актинових. Їх кількість у 15 разів менша, ніж актинових, тоді як у скелетних м’язах міозинових філаментів менше лише у 2 рази.
Наступною особливістю ГМК є відсутність тріад і погано розвинений СПР, який становить всього 2 - 7% об’єму цитоплазми.
Типи гладеньких м’язів
Розрізняють два основні типи гладеньких м’язів:
- мультиунітарний;
- моноунітарний (синцитіальний, вісцеральний).
Гладенькі м’язи мультиунітарного типу складаються з окремих, ізольованих одне від одного гладеньком’язових волокон (рис. 10.2 Б). Кожна ГМК отримує одне нервове закінчення (як і скелетне м’язове волокно). Тому одна ГМК по суті становить одну функціональну одиницю. Оскільки гладенький м’яз цього типу складається із багатьох ГМК, то він містить багато функціональних одиниць (звідки й назва – мультиунітарний).
До мультиунітарного типу належать циліарний м’яз ока, м’язи, що піднімають волосся (пілоеректори).
Гладенькі м’язи моноунітарного типу складаються із зв’язаних між собою щільними контактами ГМК (рис. 10.2 А), які скорочуються разом, утворюючи таким чином функціональний синцитій (звідки й назва – синцитіальний). До складу однієї функціональної одиниці може входити від ста до кількох мільйонів ГМК. Іннервацію отримують тільки деякі зовнішні ГМК, а через нексуси збудження поширюється на сусідні клітини.
До моноунітарного типу належать гладенькі м’язи внутрішніх органів, зокрема ГМК проток залоз, кровоносних судин, матки, сечоводів та ін.

Типи гладеньких м’язів: А- моноунітарний; Б- мультиунітарний.
Особливості функціонування
Скорочення гладеньких м’язів, як і скелетних, відбувається у 4 етапи:
1-й етап – ініціювання скорочення.
У скелетних м’язах існує лише один ініціатор скорочення - нервовий електричний імпульс. У гладеньких м’язах таких ініціаторів може бути 5:
- нервова стимуляція;
- гормональна стимуляція;
- механічне розтягнення м’язових волокон;
- зміни хімічного складу міжклітинної рідини;
- спонтанна ініціація.
Принциповою причиною для різних типів ініціації є існування на мембрані гладеньких м’язів різних типів білків-рецепторів.
Спільною рисою факторів-ініціаторів є те, що всі вони через різні механізми ведуть до збільшення внутрішньоклітинної концентрації іонів Са2+, які у гладеньких м’язах виконують 2 важливі функції: беруть участь у генерації потенціалу дії і забезпечують скорочення.
2-й етап – спряження між стимулом і скороченням.
Спряження між стимулом і скороченням досягається завдяки іонам Са2+. Основним джерелом надходження Са2+ є позаклітинне середовище. Існує великий градієнт концентрації цих іонів між позаклітинною і внутрішньоклітинною рідиною (у клітині – 10-7М, поза клітиною – 10-3М). Значення СПР як внутрішньоклітинного джерела іонів Са2+ невелике, оскільки СПР у гладеньких м’язах рудиментарний і становить усього 2 – 7% від об’єму цитоплазми.
Подальша дія Са2+ як регулятора м’язового скорочення у гладеньких м’язах істотно відрізняється від такої у скелетних м’язах. У ГМК відсутній тропоніновий механізм регуляції скорочень (немає білка тропоніну). Натомість цей механізм здійснюється іншим білком, здатним зв’язувати Са2+ - кальмодуліном. Відбувається цей процес таким чином. Іони Са2+ зв’язуються з кальмодуліном, утворюючи комплекс, який активує фермент міозинкіназу. Міозинкіназа фосфорилює (приєднує залишок фосфорної кислоти) легкі ланцюги голівок міозину. Фосфорильовані голівки міозину набувають здатності утворювати комплекси з актиновими філаментами (дефосфорильовані голівки таку здатність втрачають).
3-й етап - власне скорочення.
Молекулярні механізми скорочення такі самі, як і в скелетних м’язах,– ковзання актинових філаментів вздовж міозинових.
Основна відмінність, власне, процесу скорочення ГМК полягає у тому, що тривалість циклу утворення актоміозинового містка - його розщеплення – утворення нового містка у гладеньких м’язах значно більша (у 10 – 300 разів), ніж у волокнах скелетних м’язів. Це означає, що тривалість існування кожного, щойно утвореного актоміозинового містка у ГМК більша, як порівняти з волокнами скелетних м’язів. Причиною цього є нижча АТФ-азна активність білків. Низька швидкість утворення нових актоміозинових містків обумовлює цілу низку функціональних характеристик гладеньких м’язів. А саме:
- тривалість циклу скорочення-розслаблення у ГМК в 30 разів більша, ніж у скелетних м’язах. Гладенькі м’язи можуть перебувати у стані скорочення години і навіть дні, так званий стан тривалого м’язового тонусу;
- максимальна сила скорочення ГМК, розрахована на одиницю площі поперечного перерізу, навіть дещо більша, якщо порівняти із скелетними м’язами (гладенькі м’язи – 4-6 кг/см2, 3-4 кг/см2);
- швидкість скорочення значно менша, ніж у скелетних м’язах;
- використовують менше АТФ;
- мають малу стомлюваність.
- –й етап – розслаблення.
Цей етап пов’язаний із видаленням іонів Са2+ із клітини. Коли концентрація Са2+ зменшується до вихідного рівня 10-7, активується фермент міозинфосфатаза. Вона дефосфорилює голівки міозину, які втрачають здатність взаємодіяти з актином.
Існують такі механізми видалення Са2+ із ГМК:
- найбільше значення має робота Са2+- насоса плазматичної мембрани, який видаляє Ca2+ у позаклітинне середовище;
- Na+ - Ca2+- антипорт;
- зв’язування Са2+ зі специфічними білками СПР.
Date: 2016-11-17; view: 1137; Нарушение авторских прав