Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Происхождение жизни с точки зрения химии и геохимии
|
|
Происхождение жизни сделалось научной проблемой с тех пор, как Луи Пастер продемонстрировал невозможность самопроизвольного зарождения живых форм. По удивительному совпадению, результаты его экспериментов были опубликованы в 1859 году, в тот же год, когда Дарвин издал «Происхождение видов…», где, в числе других основополагающих идей, предложил и гипотезу об общем предке всех живых организмов. По-видимому, первое последовательное представление о происхождении жизни было изложено в 1924 году русским биохимиком Александром Ивановичем Опариным в виде популярной брошюры (Oparin, 1924), впоследствии многократно переизданной в постоянно расширяемых вариантах (Oparin and Fesenkov, 1956). Сценарий Опарина был наивным и мало обоснованным (при менее доброжелательном отношении его даже можно назвать химически несостоятельным), но он включал некоторые ключевые положения, которые принимаются в области происхождения жизни и по сей день[130]. Центральное предположение Опарина состояло в том, что среда, в которой возникла жизнь (предположительно примитивный океан в целом, но, возможно, и какая-то разновидность дарвиновского «теплого пруда»), представляла собой сложный раствор абиогенных органических молекул, включая аминокислоты и сахара, – все мономеры, необходимые для синтеза биополимеров. Опарин назвал эту гипотетическую среду, породившую жизнь, первичным бульоном (или супом). Подобный сценарий возникновения жизни был предложен позже в статье Дж. Б. С. Холдейна, уже цитированной в главе 11 по поводу гипотезы о «вирусной» стадии, предшествовавшей появлению первых клеток (Haldane, 1928). Опарин и Холдейн утверждали, что атмосфера первичной Земли была восстановительной и что первыми организмами были анаэробные гетеротрофы, питавшиеся смесью мономеров, изобиловавших в гипотетическом первичном бульоне.
Опарин и другие ранние исследователи происхождения жизни осознавали важность появления клеточной (или подобной) организации на ранней стадии жизни или даже до появления первых bona fide форм жизни. Согласно популярной тогда идее, доклеточная эволюция началась с так называемых коацерватных капель, образовавшихся в результате взаимодействия между определенными противоположно заряженными полимерами. Опарин и его соратники и последователи были биохимиками по профессии и, таким образом, отдавали в ходе своих размышлений приоритет обмену веществ. Ранние сценарии происхождения жизни придерживались того, что в коацерватах возникла простая сеть (до)метаболических реакций, обеспечивавшая обратную связь процессам роста и, возможно, деления этих пузырьков. С этого момента развивался все более сложный метаболизм, приведший в конечном итоге к появлению автотрофии. В этих качественных моделях доклеточной эволюции не уделялось никакого серьезного внимания происхождению генетической информации и процессов информационного обмена. Эти процессы (не вполне еще понятые к тому времени), как предполагалось, каким-то образом развились как следствие или побочный продукт эволюции обмена веществ. Некоторые ограниченные опыты с коацерватными каплями и другими подобными пузырьками, такими как так называемые микросферы, состоящие из беспорядочных полимеров аминокислот, известных под названием протеиноидов, продемонстрировали способность этих пузырьков поддерживать сеть простых реакций, расти и делиться, но не более того (Fox, 1976).
Не следует относиться к ранним гипотезам о происхождении жизни слишком пренебрежительно. Создатели этих качественных моделей были полностью рациональны в своем мышлении и осознавали важность обмена веществ, источников энергии и деления среды на ячейки, напоминающие клетки. Тем не менее они крайне недооценили или даже просто проигнорировали полную практическую невозможность существования «первичного бульона» в объеме всего древнего океана. Любой сколь-нибудь реалистичный сценарий происхождения жизни должен включать четко определенную доклеточную, абиогенную ячеистую структуру, неорганические катализаторы для ускорения «предбиохимических» реакций, предшествовавших появлению настоящих ферментов, градиенты температуры и (или) электрохимического потенциала, необходимые для выработки энергии в доступной форме, и решение чрезвычайно сложной проблемы о происхождения генетической информации (см. обсуждение выше в этой главе). В общем, ранние концепции недооценивали масштаб проблемы происхождения жизни и не исследовали специфические абиогенные условия, необходимые в качестве предпосылки начала биологической эволюции. Впоследствии несколько групп исследователей попытались уйти от понятия однородного первичного супа, заменив его той или иной формой неорганических ячеек, и стремились обратиться к проблеме происхождении жизни в целом, сочетая моделирование, эксперимент и наблюдения в природных условиях. Общей идеей этих гипотез является наличие единой структуры, которая может одновременно обеспечивать компартментализацию, градиенты энергии и катализаторы. Мы не можем здесь детально обсудить все эти исследования, не говоря уже обо всей области пребиотической химии. Вместо этого мы сосредоточимся лишь на нескольких моделях, представляющихся особенно продуктивными.
Г. Вахтершаузер предположил, что жизнь возникла в зонах вулканической активности, на поверхности, богатой железом, никелем, кобальтом и другими переходными металлами, каталитически активными и способствовавшими фиксации СО2, что приводило к росту органических сверхструктур. Были получены некоторые многообещающие результаты по синтезу пептидов в древних условиях, постулируемых в этой модели (Huber et al., 2003; Wächtershäuser, 1997). Таким образом, еще до того, как возникли организмы, могла существовать некая форма неорганической хемоаутотрофии. Синтезированные органические молекулы могли способствовать реакциям, катализируемым неорганическими катализаторами, приводя к химическому отбору, предвещавшему отбор биологический. Клеточная организация и генетические механизмы, согласно этому сценарию, появились в результате первичной химической эволюции.
Как уже говорилось в главе 11, Майкл Рассел и коллеги разработали, по всей вероятности, наиболее реалистичные, последовательные геохимические основы для возникновения жизни и доклеточной эволюции (Martin et al., 2008; Russell, 2007; Russell and Hall, 1997), опираясь на предшествующие исследования Дж. Бароса (Baross and Hoffman, 1985). Основная идея заключается в том, что оптимальные условия для возникновения жизни обеспечивались теплыми (но не горячими) щелочными источниками на дне первичного океана. Согласно Расселу и Холлу, гидротермальные источники образуют непрерывно-проточные реакторы, производящие насыпи осажденных карбонатов, кремния, глины и железо-никелевых сульфидов (см. рис. 12-5). Первоначально Рассел и Холл предсказали существование и размеры таких насыпей из геохимических соображений (Russell and Hall, 1997). Это предсказание было блестяще подтверждено открытием в области Срединно-Атлантического хребта образования, получившего название Потерянного города (Lost City) из-за украшающих его гигантских карбонатных шпилей (Kelley et al., 2005). Структура этих шпилей поистине замечательна: по сути, они являются сетью неорганических ячеек, образованных прежде всего сульфидами железа-никеля, способных катализировать различные органические реакции и поддерживающих постоянный поток протонов (протонодвижущая сила), который может обеспечивать энергией идущие в ячейках реакции. Как уже говорилось в главе 11, сети неорганических ячеек могли быть инкубатором жизни, от простейших реакций органического синтеза до появления и выхода из них первых клеток.
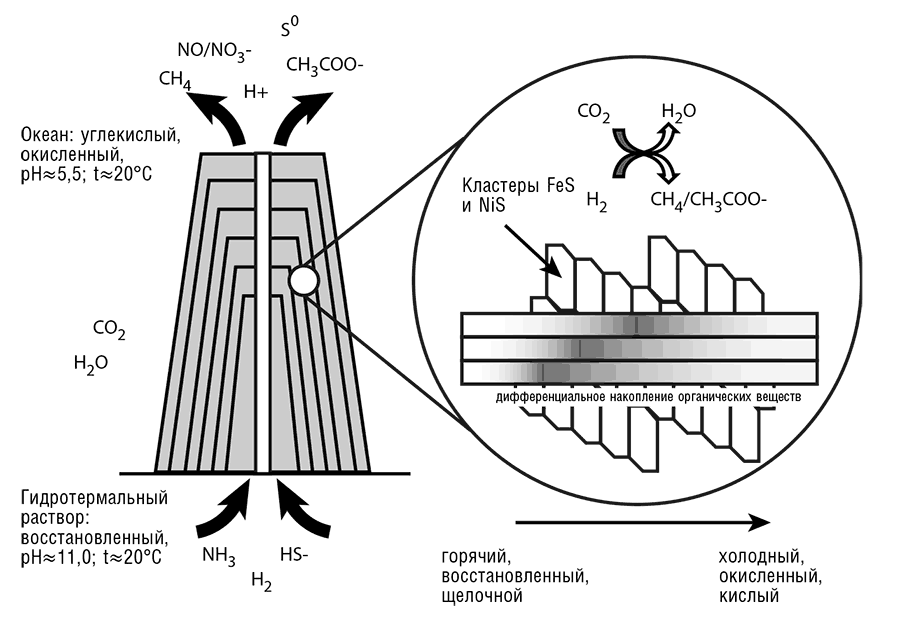
Рис. 12-5. Сети неорганических ячеек: проточные реакторы первичной химии и биохимии. В основном по данным Martin and Russell, 2007.
С целью анализа свойств этих сетей и составляющих их ячеек было проведено значительное число экспериментов и построено множество математических моделей. Было показано, что мембраны сетей полупроницаемы, так что сложные органические молекулы, включая азотистые основания, аминокислоты, сахара и жирные кислоты, оказываются изолированными в ячейках, в то время как небольшие органические молекулы и радикалы, такие как ацетат и метан, свободно сквозь них диффундируют (Mielke et al., 2010). Таким образом, ячейки представляются идеальной средой для различных реакций органического синтеза, от самых простых и фундаментальных, таких как восстановление CO2 молекулярным водородом с получением ацетата, до сложных реакций, например синтеза полинуклеотидов и пептидов, – иными словами, «происхождения биохимии» (Martin and Russell, 2007). Опытов по химии неорганических сетей пока проведено недостаточно, но компьютерное моделирование показывает, что при наличии температурного градиента, неизбежно представленного в гидротермальных источниках, может быть достигнута чрезвычайно высокая концентрация малых молекул и полимеров (Baaske et al., 2007). Такая концентрация органических молекул способна существенно облегчить различные реакции, маловероятные в ином случае, включая полимеризацию нуклеотидов и лигирование РНК (Koonin, 2007c). Хотя мембраны неорганических ячеек (Рассел заходит столь далеко, что называет их протоклетками), по всей видимости, непроницаемы для биологически важных мономеров, сами эти мембраны относительно неустойчивы (словами Рассела, «хлипкие мембраны»), так что ячейки должны разрушаться и соединяться вновь, распространяя свое содержимое. Как мы уже говорили в главе 11, в результате, вероятно, происходит отбор на уровне ячеек.
При изучении событий, случившихся лишь однажды, притом около 4 миллиардов лет назад, важно не держаться одной-единственной модели, а, так сказать, распределять свои ставки. Армен Мулкиджанян предложил существенно отличную модель неорганических ячеек, приписав роль инкубаторов самых ранних форм жизни другому виду отложений, пребывающих на меньших глубинах и при более низких температурах и состоящих в основном из сульфида цинка (Mulkidjanian, 2009). В этой модели источником энергии органических реакций является ультрафиолетовое излучение, достигающее тех небольших глубин, где находятся отложения сульфида цинка. Кроме того, сульфид цинка значительно превосходит сульфид железа в качестве катализатора и в то же время является менее реактивным химическим веществом, не оказывающим разрушительного влияния на такие лабильные молекулы, как РНК[131].
Несмотря на то что модели Вахтершаузера, Бароса, Рассела и Мулкиджаняна отличаются существенными аспектами, они объединены общими чертами, более важными, чем их различия. В каждой из этих моделей жизнь образовалась в особых местах обитания и в особых условиях, в которых неорганические ячейки способствуют накоплению органических молекул в высокой концентрации, обладают каталитическими поверхностями и обеспечивают градиент усвояемой энергии, способной поддерживать первичный органический синтез. Эти общие признаки, вероятно, должны быть частью любого стремящегося к реалистичности сценария происхождения жизни.
Радикальная альтернатива: космология вечной инфляции, переход от случайности к биологической эволюции в истории жизни и переоценка роли крайне редких событий в эволюции [132]
Как уже отмечалось выше, ситуация в области происхождения жизни в целом видится довольно мрачной. Даже при (весьма нетривиальном) предположении, что мономеры, такие как НТФ, изобильны и доступны, проблема синтеза достаточно стабильных полимеров с регулярной структурой (РНК) чудовищно сложна, а проблема происхождения репликации и трансляции таких первоначальных молекул РНК еще сложнее. В этой книге неоднократно подчеркивалось, что эволюция путем естественного отбора и дрейфа может начаться только после возникновения достаточно точной репликации, причем даже на этой стадии эволюция трансляции остается весьма проблематичной.

Рис. 12-6. Добиологическая и биологическая стадии происхождения жизни: переход от антропной причинности к биологической эволюции.
Появлению первой репликаторной системы, ознаменовавшему собой «дарвиновский прорыв»[133], неизбежно предшествовала череда сложных, трудных шагов, на которых биологические эволюционные механизмы еще не действовали (см. рис. 12-6). Даже с учетом среды, способной облегчить эти процессы, такой как сети неорганических ячеек в гидротермальных источниках, умножение вероятностей этих шагов делает появление первых репликаторов ошеломляюще невероятным (см. прил. II).
Глубокие трудности проблемы происхождения жизни могут показаться непреодолимыми, что заставляет задаться весьма общими вопросами, выходящими за рамки биологии. Не существует ли скрытых от нас факторов, имеющих решающее значение для зарождения жизни, которые, будь они нам сейчас известны, существенно изменили бы картину и сделали бы возникновение жизни гораздо более вероятным? Возможно ли, что процессы, формирующие основу для возникновения жизни, действительно так сложны, как мы себе представляем, но число попыток настолько велико, что появление жизни в результате одной или нескольких из них возможно или даже неизбежно? Другими словами, возможно ли, что сами наши понятия о вероятности неполны?
Первая возможность имеет отношение к нахождению условий, существовавших на первичной Земле и каким-то образом сделавших происхождение жизни «легким». Ячейки Рассела продвигают нас на этом пути, но, видимо, недостаточно далеко: даже в подобных проточных реакторах, богатых энергией и катализаторами, сочетание всех необходимых процессов было бы крайне редким.
Вторая возможность может быть рассмотрена в контексте всей Вселенной. Зададимся вопросом, сколько существует планет с благоприятными условиями для возникновения жизни, иными словами, сколько попыток возникновения жизни всего было сделано. В данном разделе мы рассмотрим эту линию рассуждений с точки зрения современной физической космологии.
В XX веке космология прошла драматический путь от странного (и не особенно авторитетного) философского направления к полноценному, быстро развивающемуся разделу физики, прочно опирающемуся на многочисленные научные наблюдения. Ведущее направление в сегодняшней космологии сосредоточивается на так называемой инфляции, периоде экспоненциально быстрого первоначального расширения Вселенной (Carroll, 2010; Guth, 1998a; Guth and Kaiser, 2005; Vilenkin, 2007). В наиболее правдоподобных, самосогласованных моделях инфляция вечна, с бесконечным числом островных вселенных (или просто вселенных), возникающих через распад небольших регионов изначального океана ложного (высокоэнергетического) вакуума и составляющих бесконечную мультивселенную (см. прил. II). Модель «мира многих миров» (МММ) делает поразительное предсказание, что все макроскопические, «крупнозернистые» истории событий, не запрещаемых физическими законами сохранения, были реализованы (или, возможно, будут реализованы) где-то в бесконечной мультивселенной, и не только однажды, но бесконечное число раз (Garriga and Vilenkin, 2001; Vilenkin, 2007). Например, существует бесконечное число (макроскопически) точных копий Земли, со всем, что существует на ней, хотя вероятность того, что данная наблюдаемая область Вселенной содержит одну из этих копий, исчезающе мала. Эта картина выглядит крайне нелогичной, и даже сумасбродной, но таково прямое следствие вечной инфляции, доминирующей модели эволюции Вселенной в современной космологии.
Модель МММ тесно связана с антропным принципом (иногда называемым антропным отбором), спорной, но мощной и популярной среди космологов концепцией (Barrow and Tipler, 1988; Carter, 1974; Livio and Rees, 2005). Согласно антропному принципу, единственная причина, по которой наша Вселенная имеет свои характерные параметры, состоит в том, что в противном случае в этой Вселенной не было бы никаких наблюдателей[134]. В такой форме так называемый «слабый» антропный принцип может быть реалистично определен только в контексте огромной (или, еще лучше, бесконечной) мультивселенной. В модели МММ антропный отбор имеет простой смысл: среди огромного числа наборов параметров, существующих в мультивселенной (в бесконечном числе экземпляров каждый), наша Вселенная должна иметь именно такие параметры, которые способствуют возникновению и поддержанию сложных форм жизни. Иногда говорят, что наша Вселенная принадлежит «биофильной области» мультивселенной (Livio and Rees, 2005). Сам термин «антропный принцип» можно считать неудачным, так как он может быть истолкован как предполагающий некую особую важность людей или, в более общем случае, сознательных наблюдателей или, что еще хуже, привлечь телеологические интерпретации. Такой взгляд на антропный принцип совершенно неверен; на самом деле он представляет собой просто «отбор наблюдателя» (Bostrom, 2002). Тот факт, что в этой Вселенной имеется жизнь, строго ограничивает ее характеристики: по меньшей мере, наша часть Вселенной должна содержать галактики и планетные системы, а не только массивные черные дыры или частицы разреженного газа, являющиеся в принципе более вероятными состояниями с высокой энтропией.
В сравнении со старыми космологическими концепциями, рассматривавшими конечную Вселенную, модель МММ меняет самые определения возможного, вероятного и случайного по отношению к любому историческому сценарию. Проще говоря, вероятность реализации любого сценария, не запрещенного законами сохранения, в бесконечной мультивселенной в точности равна единице. С другой стороны, вероятность того, что данный сценарий реализуется в данной (островной) вселенной, равна частоте этого сценария в мультивселенной и может быть исчезающе малой. С несколько иной точки зрения известная идея второго начала термодинамики, будучи истинной лишь в статистическом смысле, обретает буквальный смысл в бесконечной мультивселенной: любое нарушение второго начала, позволенное законами сохранения, действительно случилось, причем бесконечное число раз. Таким образом, спонтанное возникновение сложных систем, которое могло бы считаться практически невозможным в конечной вселенной, становится не только возможным, но и неизбежным в МММ, хотя априорная вероятность подавляющего большинства историй, происходящих в данной вселенной, исчезающе мала. Эта новая сила случайности, подкрепленная антропным рассуждением, имеет глубокие последствия для нашего понимания любого явления во Вселенной, не исключая и жизнь на Земле (Koonin, 2007b).
История жизни не может не включать в себя важнейший переход от случая к биологической эволюции (см. рис. 12-6). Синтез нуклеотидов и полинуклеотидов по крайней мере среднего размера не мог возникнуть биологически и, следовательно, произошел абиогенным путем, то есть фактически случайно, при помощи химического отбора, как, например, выживание более стабильных видов РНК. На другом конце спектра не может быть никаких разумных сомнений в том, что первые клетки произошли в результате биологической эволюции на ее доклеточной стадии (см. гл. 11). Где-то между этими крайностями имеется переход, порог биологической эволюции. Чаще всего, с появлением концепции мира РНК, этот порог неявно связывается с появлением репликации молекул РНК. Трансляция, как полагают, возникла позже, в результате специфического процесса, движимого отбором. Как уже говорилось в предыдущем разделе, как катализируемая рибозимами репликация, так и, особенно, эволюция трансляции в мире РНК сталкиваются с огромными трудностями. Модель МММ значительно расширяет интервал на оси организационной сложности, где порог может быть преодолен через появление сложности, возникшей случайно (см. рис. 12-6). В этой модели возможность того, что прорывная стадия наступления биологической эволюции была состоянием высокой сложности, не может быть сброшена со счетов, какой бы невероятной (то есть редкой в смысле мультивселенной) и нелогичной она ни казалась. Например, в рамках этой модели прорыв может быть обеспечен случайным появлением центральных элементов сопряженной системы репликации и трансляции, напоминающей, по крайней мере в принципе, современные РНК-вирусы (Koonin, 2007b).
Модель МММ не только допускает, но и гарантирует, что где-то в бесконечной мультивселенной (более того, в каждой отдельной бесконечной вселенной) такая сложная система появится; кроме того, существует бесконечное число этих систем. Таким образом, уместный вопрос состоит не в том, возникла ли система произвольной сложности спонтанно, по воле случая (поскольку МММ гарантирует это), но в том, каков был наиболее вероятный прорывный этап на Земле, который должен быть отнесен к случайности в соответствии с антропным подходом. Я полагаю, что, учитывая серьезные проблемы эволюционных сценариев, разработанных, чтобы объяснить происхождение репликации и трансляции путем биологической эволюции, следует принимать всерьез возможность того, что порог биологической эволюции соответствует очень сложной стадии – возможно, системе репликации вкупе с трансляцией и с белковыми полимеразами, отвечающими за репликацию РНК. Эта гипотеза (которую я называю антропной химической эволюцией, АХЭ), конечно, не исключает особой важности рибозимов в ранней биологии, в частности, в первичной системе трансляции, как предполагается исходя из сравнительного анализа белковых компонентов аппарата трансляции (см. выше в этой главе). Тем не менее следствием из модели АХЭ является то, что мир РНК, каким он описывается в настоящее время, – огромное сообщество реплицирующихся молекул РНК, наделенных различными каталитическими возможностями, но не содержащий ни системы трансляции, ни генетически кодируемых белков, – мог никогда и не существовать.
Согласно гипотезе АХЭ, основные элементы системы трансляции, а именно состоящая только из РНК рибосома и избирательные адаптеры, по крайней мере для некоего подмножества из двадцати современных белковых аминокислот, возникли случайно, в соответствии с антропным принципом. Согласно этой модели, прорывная система, запустившая биологическую эволюцию, была примитивной, но достаточно эффективной РНК-машиной трансляции, способной транслировать экзогенную РНК, генерируя функциональные белки, в том числе репликазы. Достаточное разнообразие случайно синтезированных РНК, включая и те, что кодировали белки, обладающие активностью репликазы (хоть первоначально и низкой), является еще одной антропно определенной особенностью тех мест на древней Земле, где зародилась жизнь. Как уже говорилось в предыдущем разделе, сети неорганических ячеек в гидротермальных источниках могли играть роль добиотических химических реакторов. Существование такой сети само по себе также является частью антропного сценария.
В таких условиях появление основанного на РНК механизма трансляции привело бы к производству репликазы и, с последующей репликацией РНК, к критическому переходу от антропной причинности к биологическому отбору (см. рис. 12-6). В принципе начало биологической эволюции можно себе представить и с репликазой как единственным изначально активным белком. Однако, учитывая вероятное существование реакторов, производящих РНК, которые обсуждались ранее в этой главе, вполне возможно, что с появлением трансляции другие случайные последовательности РНК послужили основой предковых форм наиболее распространенных белковых укладок, порождая несколько белковых функций (например, РНК-связывающие белки и примитивные ферменты, катализирующие синтез нуклеотидов) и тем самым придавая минимально необходимую устойчивость формирующейся биологической системе. Появление этих укладок можно назвать Большим взрывом белковой эволюции.
Как уже отмечалось, современный универсальный генетический код гораздо более надежен, чем был бы случайный, по отношению к мутационным и, вероятно также, к трансляционным ошибкам. Эта устойчивость проявляется и в очевидной неслучайности структуры кода, выражающейся в первую очередь в том, что серия кодонов, которые отличаются только третьей позицией, кодирует либо одну и ту же, либо две подобные аминокислоты, и в других особенностях соответствия кодонов аминокислотам (Koonin and Novozhilov, 2009). Примечательно, что предполагаемый предковый «дублетный» код, в котором третья позиция не несла никакой информации, мог быть даже более надежным, чем современный (Novozhilov and Koonin, 2009). Надежность, как обычно предполагается, эволюционировала в ходе оптимизации кода. Однако модель АХЭ предлагает альтернативную точку зрения, при которой базовая структура кода возникла по чистой случайности, поскольку только коды с определенным минимальным уровнем надежности позволили бы функциональной репликазе появиться в прорывной системе. Конечно, этот сценарий не исключает и последующей корректировки кода биологической эволюцией, что, по всей вероятности, в действительности произошло.
Таким образом, гипотеза АХЭ снимает парадоксы происхождения репликации и трансляции предположением о том, что оба этих процесса, в их примитивных формах, не произошли в результате биологической эволюции, а возникли случайно, как связанная система, в соответствии с антропным принципом.
Гипотеза АХЭ, несомненно, должна показаться большинству эволюционных биологов нелепой и возмутительной, поскольку она уклоняется от поисков «механизмов» доклеточной эволюции. Тем не менее существуют смягчающие обстоятельства. Во-первых, постулируемая возможность возникновения сопряженной репликационно-трансляционной системы не требует никаких неизвестных процессов. Напротив, для нее необходимы только хорошо известные, обычные реакции, такие как полимеризация нуклеотидов и аминокислот и фосфорилирование/дефосфорилирование нуклеотидов; при этом нужны только распространенные в химии и биохимии взаимодействия. Как уже отмечалось в этой главе, элементарные реакции, необходимые для трансляции (активация аминокислот, аминоацилирование РНК и транспептидация), легко моделируются с помощью рибозимов, в противоположность репликации РНК, которой, как известно, трудно достичь в отсутствие белков. Во-вторых, исключая разве что полную неадекватность нынешнего понимания условий на первичной Земле, любые мыслимые сценарии эволюции жизни обязательно требуют сочетания маловероятных условий и событий до начала биологической эволюции. Список таких событий включает в себя абиогенный синтез довольно сложных и не очень устойчивых органических молекул, таких как нуклеотиды, накопление этих молекул в соответствующих ячейках до высоких концентраций и их полимеризацию с получением полинуклеотидов достаточного размера и разнообразия. Таким образом, независимо от космологических соображений, некоторые формы антропной причинности представляются неизбежным аспектом эволюции жизни (см. рис. 12-6).
Я привел сценарий АХЭ, чтобы показать, что диапазон сложности, открытой для антропной причинности, может быть гораздо шире, чем предполагалось ранее, настолько, что первичная репликационно-трансляционная система могла возникнуть без биологического отбора. Случайное происхождение изощренной системы, способной исполнять сложные биологические функции, может показаться бессмыслицей. Я полагаю, однако, что это всего лишь семантическая ловушка. До наступления биологической эволюции не может быть «функции», только сложность, а модель МММ гарантирует возникновение любого уровня сложности (это гарантированно произойдет «где-то» в бесконечной Вселенной, но антропный принцип прямо помещает эти события на Землю).
Все эти соображения провоцируют довольно-таки кошмарный вопрос: имеет ли какое-либо значение в бесконечно избыточном мире МММ биологическая эволюция вообще и дарвиновский отбор в частности? Разве не возникнет система любой, даже наивысшей, сложности просто случайно? Ответ: да, но этот вопрос упускает важное обстоятельство. В модели МММ случайное появление бесконечного числа сложных биот неизбежно, но несравнимо реже, чем развитие по сценарию АХЭ, которое включает в себя переход от случайности к биологической эволюции после стадии прорывной системы (см. рис. 12-6). Начало биологической эволюции канализирует исторический процесс, сокращая многочисленные траектории, которые возможны в принципе, до относительно малого числа стабильных и более вероятных – тех, что совместимы с дарвиновским режимом эволюции сложных систем (см. рис. 12-7). Этот переход приводит к гораздо большей скорости эволюционных изменений, чем та, что достигается случаем, и, как только возникает возможность для биологической эволюции, антропная причинно-следственная связь отходит на второй план в истории жизни. Конечно, «второй план» не означает потери важности: непредвиденные случайности имеют решающее значение, особенно на переходных этапах эволюции (см. обсуждение ранее в этой книге, особенно в гл. 7). Таким образом, в любой реконструкции происхождения жизни и ранней эволюции порог должен быть связан с самой нижней точкой, то есть минимально сложной системой, способной к биологической эволюции.

Рис. 12-7. Отсечение эволюционных траекторий на пороге биологической эволюции: а – только химическая эволюция; б – возникновение биологической эволюции.
Сильная форма гипотезы АХЭ, согласно которой прорывным этапом в истории жизни была примитивная совместная репликационно-трансляционная система (см. рис. 12-6), в принципе легко опровержима. Такая система должна рассматриваться как верхняя граница сложности для прорывного этапа. Как только возможность биологической эволюции при более низком уровне сложности, таком как мир РНК, будет убедительно продемонстрирована, а путь от мира РНК к системе трансляции очерчен либо опытным путем, либо в убедительной модели, сильная форма гипотезы АХЭ будет опровергнута. Демонстрация самостоятельного возникновения жизни на нескольких планетах в нашей Вселенной будет иметь тот же результат. В приложении приведен грубый, но, хочется надеяться, поучительный расчет верхней границы вероятности возникновения совместной репликационно-трансляционной системы в наблюдаемой части Вселенной; эта вероятность, несомненно, исчезающе мала. Противоположное предсказание состоит в том, что любые формы жизни, которые могут быть обнаружены на Марсе, или, возможно, Европе (спутнике Юпитера, на котором была обнаружена жидкая вода), или даже на любой экзопланете в ходе будущих планетарных исследований, будут иметь общее с земной жизнью происхождение. Любая из этих находок опровергнет сильную гипотезу АХЭ, но не сделает модель МММ незначимой для нашего понимания происхождения жизни. Действительно, любое такое открытие (важное само по себе) просто понизит порог биологической эволюции в масштабе рис. 12-7.
Самым простым и сильным опровержением гипотезы АХЭ будет опровержение самой МММ[135]. Тем не менее здесь требуется важное разъяснение. Для обоснованности представленных здесь концептуальных рамок не требуется верная во всех деталях МММ. Только два общих положения существенны: (1) пространственно бесконечная вселенная, такая, как любая островная вселенная в МММ; мультивселенная, хоть и является неотъемлемой частью теории вечной инфляции, не требуется в качестве аргумента; (2) конечность числа различных макроскопических историй. Даже сильная форма гипотезы АХЭ, представленная здесь, не будет опровергнута, если некоторые конкретные детали МММ окажутся неправильными, но только если одно из этих общих предположений окажется неверным.
Date: 2016-05-15; view: 723; Нарушение авторских прав