Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Сравнительно-геномная реконструкция генного репертуара LUCA
|
|
Почему мы уверены, что LUCA существовал? Есть множество аргументов, поддерживающих предположение о существовании LUCA, и самым сильным из них является сохранившаяся в универсальном (одинаковом для всех организмов) виде система экспрессии генов. Действительно, все известные нам клеточные формы жизни используют один и тот же генетический код (20 универсальных аминокислот и стоп-сигналов, закодированных в 64 кодонах), и небольшие отклонения содержатся лишь в сильно деградировавших геномах бактериальных паразитов и органелл. Все клетки используют одинаковые рибосомы, которые состоят из трех универсально консервативных молекул РНК и примерно 50 белков, из которых 20 универсальны. В дополнение к этому, универсально сохраняющиеся компоненты системы трансляции включают около 30 транспортных РНК, несколько факторов трансляции, 18 аминоацил-тРНК-синтетаз и несколько тРНК-модифицирующих ферментов (Anantharaman et al., 2002). За пределами системы трансляции универсально консервативны только гены трех субъединиц РНК-полимеразы. Таким образом, существует всего около 100 универсально сохранившихся генов, и все они участвуют в экспрессии генов (случается, что некоторые из этих генов, особенно те, что кодируют малые рибосомные белки, отсутствуют в аннотациях вновь расшифрованных геномов [Charlebois and Doolittle, 2004]; однако, скорее всего, эти исчезновения являются ошибками расшифровки или аннотации, а не реальными потерями в геноме). Мы уже знакомы с этими (почти) универсальными генами из главы 6, где мы наблюдали весьма сходные (хотя и не тождественные) топологии их филогенетических деревьев. Универсальное сохранение генетического кода и механизма экспрессии и единство эволюционных судеб его компонентов не оставляют места для сомнений в том, что эта система является наследством некоей формы LUCA. Основной вопрос состоит, таким образом, не в том, существовал ли LUCA, а в том, каким он был, о каких свойствах этой системы мы можем сделать выводы с достаточной уверенностью, а какие (пока что) остаются неясными[114].
Ясно, что ни один организм, даже самый примитивный, не может состоять из одних механизмов экспрессии, так что необходима реконструкция остальной части генного репертуара LUCA. И методы реконструкции, и их подводные камни здесь те же, что были описаны в главах 5 и 7. В контексте эволюции прокариот, которая важна для реконструкции LUCA, следует рассматривать три типа элементарных событий: (1) «рождение» гена – появление нового гена, обычно в процессе генной дупликации, за которой следует радикальная дивергенция; (2) обогащение генами за счет ГПГ; и (3) утрата гена. Напомним, что надежной реконструкции пути эволюции и наборов предковых генов препятствует неточность, связанная с относительными вероятностями событий и скоростями разных процессов, в особенности разница скоростей утраты генов и ГПГ. В принципе даже ген, найденный во всех современных клеточных формах жизни, может не быть унаследован от LUCA: его универсальность может быть последствием многочисленных горизонтальных переносов. Более того, предположения о генном содержании предковых форм, основанные на принципе наибольшей экономии и даже наибольшего правдоподобия, консервативны, и неясно, насколько они недооценивают результат. Вопреки всем сложностям и неточностям эволюционных реконструкций, анализ наибольшей экономии совместно с менее формальными попытками реконструкции далекого прошлого конкретных функциональных систем не оставляет серьезных сомнений в том, что LUCA уже содержал несколько сотен генов (Mirkin et al., 2003; Ouzounis et al., 2006; Snel et al., 2002). В дополнение к вышеупомянутой «золотой сотне» генов, участвующих в экспрессии, этот разнообразный генный комплект состоит из множества метаболических ферментов, включающих как пути основного энергетического обмена и биосинтеза аминокислот, нуклеотидов и коферментов, так и некоторые важные белки мембран, такие как субъединицы частицы узнавания сигналов (SRP) и H+-АТФазы.
Однако в реконструкции генного состава LUCA по-прежнему зияют дыры. Более всего удивляет (1) отсутствие ключевых компонентов механизма репликации ДНК, а именно полимераз, ответственных за инициацию (праймазы) и элонгацию репликации ДНК и заполнение лакун после удаления праймера, а также основных геликаз (Leipe et al., 1999); и (2) отсутствие основных ферментов липидного биосинтеза (Pereto et al., 2004). Эти незаменимые белки не попадают в реконструированный генный репертуар, поскольку данные процессы у бактерий и архей катализируются разными, неродственными ферментами, а в случае мембранных фосфолипидов еще и образуют химически разные мембраны[115].
Таким образом, реконструированный генный набор LUCA исключительно неоднороден: в то время как отдельные системы по своей сложности почти не отличаются от аналогичных систем существующих организмов, другие системы выглядят рудиментарными или вовсе отсутствуют. Столь странная картина напоминает концепцию асинхронной «кристаллизации» различных клеточных систем на ранних стадиях эволюции, которую предложил Карл Вёзе (Woese, 1998), и подсказывает, что нужно взглянуть на проблему LUCA с более общих позиций.
По-видимому, имеет смысл думать о LUCA в двух разных плоскостях (Koonin, 2009c):
• генетическая сложность, которая может быть выражена в числе генов;
• степень структурного и биологического сходства с современными клетками. Для краткости и удобства назовем эти свойства «клеточностью».
Можно ожидать, что эти характеристики коррелируют между собой, но они не связаны жестко. В принципе вполне можно представить, что LUCA был клеточной формой жизни значительно более простой, нежели любая из современных клеток (по крайней мере свободноживущих) в смысле генетического содержания; или же, напротив, что относительная генетическая сложность была свойственна клеткам еще до появления современного типа клеточной организации – для этого сценария мы будем использовать обозначение LUCAS (Last Ancestral Universal Common State – последнее универсальное предковое состояние [всего живого]; см. рис. 11-1).
«Униформистское предположение» о том, что LUCA был более-менее обычной, похожей на современные прокариотической клеткой, негласно принимается в большинстве описаний ранней клеточной эволюции, но редко проговаривается явно. Тем не менее любая реконструкция LUCA должна учитывать и эволюцию тех свойств, которые трудно проследить вглубь до единого предка архей и бактерий, особенно два главных из них: репликацию ДНК и биогенез (и химию) мембран. Униформистские гипотезы, которые скрыто или явно основаны на предположении о «клеточности» LUCA, объясняют несохранение этих ключевых систем (на самом глубоком уровне реконструкции) одним из двух способов:
• каким-то образом LUCA комбинировал разные варианты этих систем, a впоследствии в архейной и бактериальной линиях эволюции был утерян один из вариантов, свой для каждой линии;
• LUCA обладал определенным вариантом каждой из этих систем, впоследствии замененным на неортологичный у архей или бактерий.
Точнее, говоря о биогенезе мембран, предполагают, что LUCA имел смешанную гетерохиральную мембрану, а две хиральные разновидности появились в результате последующей специализации архей и бактерий соответственно (Pereto et al., 2004). Касательно же репликации ДНК была высказана гипотеза о том, что одна из современных систем репликации является предковой, в то время как другая развивалась у вирусов и впоследствии заменила собою исходную либо у архей, либо у бактерий (Forterre, 1999, 2006).
В противоположность этому, более радикальные предположения относительно природы LUCA исходят из положения «что наблюдаемо, то и есть»: LUCA вообще не обладал этими ключевыми чертами, общими для бактерий и архей, по крайней мере не обладал ими в их современной форме (Koonin, 2009c; Koonin and Martin, 2005). Возможность того, что LUCA существенно отличался от всех известных ныне клеток, была впервые высказана в концепции «прогеноты» – гипотетической примитивной формы жизни, в которой связь между генотипом и фенотипом еще не полностью сформировалась (Doolittle and Brown, 1994; Woese and Fox, 1977). В своей первоначальной форме идея прогеноты включает примитивную неточную трансляцию – не слишком жизнеспособное предположение, с учетом обширной диверсификации белков до LUCA, которую однозначно демонстрирует анализ различных белковых семейств (см. гл. 12).
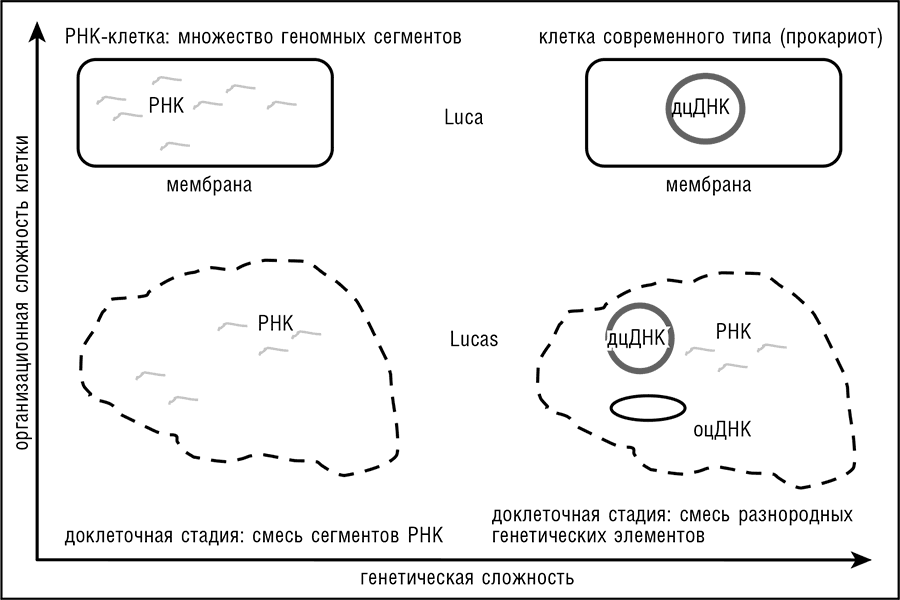
Рис. 11-1. Пространство логических возможностей для LUCA(S): генетически сложный или генетически простой, клеточный или неклеточный. Предполагается, что доклеточный пул молекул РНК генетически проще, чем смешанный пул различных генетических элементов (LUCAS), и что предполагаемая РНК-клетка проще, чем клетки современного типа (LUCA).
Более реалистично предполагать, что развитие основных черт клетки («кристаллизация» в смысле Вёзе) была асинхронной, так что LUCA сильно напоминал современные клетки в одних аспектах, но был исключительно «примитивен» в других. Результаты сравнительной геномики дают подсказки для различения «развитых» и примитивных свойств LUCA. Так, основываясь на основных областях несходства между археями и бактериями, была высказана гипотеза о том, что LUCA
• не имел типичного большого ДНК-генома;
• не был типичной, окруженной мембраной клеткой (см. рис. 11-2; Koonin and Martin, 2005).
Загадкой ДНК-генома и репликации, которую требовалось разгадать, была комбинация негомологичных и консервативных компонентов в аппаратах репликации ДНК архей и бактерий, притом что ядро механизма транскрипции универсально консервативно. Чтобы объяснить эту смешанную картину консерватизма и разнообразия, было сделано предположение о том, что у LUCA был «ретровирусоподобный» цикл репликации с консервативным механизмом транскрипции, вовлеченным в транскрипцию дцДНК провирусного типа и консервативными компонентами системы репликации ДНК, которые в данном случае играли вспомогательную роль (Leipe et al., 1999). Эта умозрительная схема сочетала в некоем гипотетическом цикле репликации консервативные белки, вовлеченные в транскрипцию и репликацию, с белками, которые, по крайней мере в современной биосфере, однозначно принадлежат к миру вирусов, как, например, обратная транскриптаза (см. гл. 10). Этот предполагаемый изначальный цикл репликации-экспрессии, хотя формально и подходит под требования универсального сохранения упомянутых белков при неуниверсальности других ключевых компонент механизма репликации ДНК, не имеет, однако, никаких аналогий в существующих генетических системах.
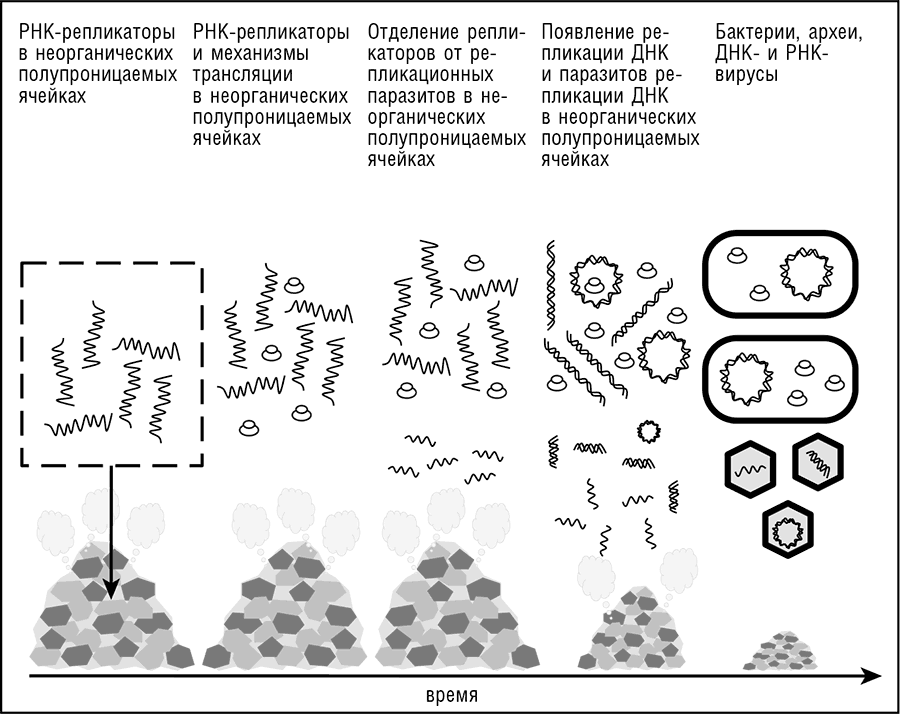
Рис. 11-2. Сценарий вирусного мира для доклеточной эволюции.
Еще одна область несходства архей и бактерий, липидный биосинтез (вместе с химией липидов), требует еще более радикальной гипотезы неклеточного, но компартментализированного LUCA (Martin and Russell, 2003; Koonin and Martin, 2005). Конкретное предположение состоит в том, что LUCA(S) был разнородной популяцией генетических элементов, существовавших в сети неорганических ячеек (в гл. 12 эти возможные инкубаторы жизни обсуждаются в подробностях). Основным затруднением для моделей безмембранного LUCA является то, что некоторые мембранные белки и даже молекулярные комплексы, такие как протонная АТФаза и частица узнавания сигнала, практически универсальны во всех современных формах клеточной жизни и, по всей вероятности, присутствовали и в LUCA.
Более тщательное рассмотрение основные проблем концепции клеточного LUCA, а именно негомологичности основных элементов системы репликации ДНК и радикального различия между фосфолипидами и ферментами липидного биосинтеза у архей и бактерий, приводит к идее о том, что две эти проблемы тесно связаны между собой (Koonin, 2009c). LUCA (реконструированный методами сравнительной геномики) как сложный организм без обширного ДНК-генома, сравнимого с геномами современных архей и бактерий, должен был бы иметь геном, состоящий из нескольких сотен РНК-сегментов (или ДНК провирусного типа), длиной в несколько тысяч оснований каждый. Это ограничение диктуется исключительно низкой стабильностью молекул РНК в сравнении с ДНК, а также тем фактом, что самые длинные РНК-геномы (принадлежащие коронавирусам) не превышают 30 Кб.
Предполагалось, что LUCA мог быть РНК-клеткой в собственном смысле слова, из которой далее произошли три основные линии РНК-клеток (предки бактерий, архей и эукариот), геномы которых независимо заменились ДНК в результате приобретения механизмов ДНК-репликации от различных вирусов (Forterre, 2006). Этот сценарий, однако, выглядит надуманным, поскольку необходимость в сотнях «хромосом» выглядит непреодолимым препятствием для РНК-клетки. Требуемая точность распределения копий такого фрагментированного генома между дочерними клетками при клеточном делении требует исключительной точности механизма сегрегации генома, превышающей даже точность соответствующего аппарата современных прокариот. В противном случае изменение в наборе генов при каждом делении клетки, безусловно, сделает размножение невозможным. Механизмы деления, которые функционируют в современных бактериях (и, вероятно, в археях), закачивают дцДНК в дочерние клетки при помощи специальных АТФаз, которые, по всей вероятности, эволюционировали совместно с дцДНК-геномами. Таким образом, если LUCA действительно имел, вместо большого генома из дцДНК, «коллективный» геном из множества РНК-сегментов, то он должен был быть формой жизни, весьма отличной от современных клеток, возможно даже неклеточной.
Другим широко обсуждаемым аспектом ранних форм жизни, включающих и LUCA, является безудержный ГПГ, который часто рассматривается как предпосылка к эволюции сложной жизни (еще одно важное предположение Карла Вёзе [Woese, 2002]). Несомненно, ГПГ – путь к быстрым новоприобретениям, а на ранних стадиях эволюции жизни новоприобретения должны были быть исключительно быстрыми. Вёзе и его коллеги предположили и проиллюстрировали на математических моделях, что сама универсальность генетического кода могла быть связана с критической ролью ГПГ на ранних стадиях эволюции (Vetsigian et al., 2006). При обширном ГПГ единый вариант кода обязательно распространится во всей популяции предковых форм, поскольку любой организм с отличающимся генетическим кодом будет неспособен получать выгоду от ГПГ и, будучи изолированным от других организмов, непременно будет отбракован отбором[116]. Постоянный бурный ГПГ – неотъемлемая черта моделей неклеточного компартментализированного LUCA(S), но, безусловно, не может быть принят как данность в рамках моделей клеточного LUCA. В следующем разделе мы рассмотрим модель неклеточного LUCA(S) подробнее.
Date: 2016-05-15; view: 755; Нарушение авторских прав