Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Генетические исследования условной реакции активного избегания
|
|
Активное избегание удара электрического тока в челночной камере (см. гл. 3)— это четкий тест на обучаемость у лабораторных грызунов, показатели которого легко измерить. Он достаточно унифицирован и
его результаты, полученные в разных лабораториях, легко сопоставить друг с другом. Поскольку в качестве условного раздражителя можно выбрать звук, то тест позволяет включать в сравнение и животных-альбиносов, не опасаясь, что свойственная им низкая острота зрения повлияет на успешность выполнения навыка.
В начале 60-х годов на основе популяции крыс Вистар итальянский исследователь Дж. Биньями начал селекцию на высокую и низкую способность крыс к обучению реакции активного избегания в челночной камере. Уже через несколько лет между селектированными линиями существовали достоверные различия в обучаемости: линии были сформированы. Они получили название Римских (Roman High Avoidance, RHA, Roman Low Avoidance, RLA). С начала 70-х годов селекция и разведение этих крыс проводились независимо в нескольких лабораториях разных стран (Driscoll, Battig, 1982; Fernandez-Teruel et al., 1997).
Исследование крыс Римских линий с помощью практически всех существующих методик оценки поведения дало основание считать, что наиболее сильные межлинейные различия у RHA и RLA связаны с эмоциональностью и разным типом реакции крыс двух линий на стрессоры.
Эти линии тем не менее все же различаются и по способности к ассоциативному обучению как таковой.
В 70-е годы путем селекции были созданы еще две линии крыс — Сиракузские (Syracuse High Avoidance, SHA, Syracuse Low Avoidance, SLA). Они, как и Римские линии, достоверно различались по скорости усвоения реакции активного избегания, но (в отличие от Римских), не различались по уровню двигательной активности. Исходной популяцией для этой селекционной работы послужили крысы Лонг-Шанс. В настоящее время крысы хорошо обучающейся линии 577Адают примерно 40 реакций избегания в 60 предъявлениях теста, тогда как SLA — ни одной. У плохо обучающейся линии SLA было мало межсигнальных реакций, однако скорость выполнения реакций избавления (т.е. реакции на включение собственно болевого стимула — электрического тока) у них не отличалась от линии SHA. Иными словами, межлинейные различия обнаруживались в «готовности» к выполнению перехода из одной половины камеры в другую, но не затрагивали реакции на боль. В тесте «открытого поля» у Сиракузских, так же как у Римских линий, различался уровень возбудимости вегетативной нервной системы. Речь идет о так называемой «эмоциональности» крыс, которая оценивается по числу болюсов дефекации при помещении животного в новую, слегка пугающую обстановку теста «открытого поля». SLA, так же как и RLA, оказались более «эмоциональными», т.е. испуг у них был сильнее.
В Возможно, что менее эффективное обучение реакции избегания, общее для линий RLA и SLA, имеет одну причину — повы-
В" шенную пугливость этих животных, которая препятствует образованию ассоциации между условным сигналом и реакцией животного.
Различия в процессах обучения и памяти в связи с генетической изменчивостью строения мозга. В соответствии с традиционно принятой в нейрофизиологии логикой исследований функциональную роль того или иного отдела мозга в формировании поведения обычно анализировали путем оценки последствий его разрушения, а также электрической и/или фармакологической стимуляции.
К началу 70-х годов считалось установленным, что одна из функций гиппокампа (рис. 9.2А) — мощное модулирующее влияние на процессы обучения, в частности торможение инструментальных ус-ловнорефлекторных реакций (Виноградова, 1975).
Американские исследователи Р. и Ц. Ваймеры и Т. Родерик выполнили исследование, в котором анализировалась роль генотипических особенностей в обеспечении функции гиппокампа. Способность мышей генетически гетерогенной популяции к обучению пассивной реакции избегания удара электрического тока (при однократном его применении) авторы сопоставили с общим объемом гиппокампа, который определяли после окончания экспериментов. Для этого на срезах мозга каждого животного, прошедшего тест на обучение, определили площадь, занимаемую гиппокампом, а затем в соответствии с существующими морфометрическими правилами вычислили его суммарный объем (Wimer et al., 1971).
в Сопоставление результатов опытов с поведением и данными | подсчетов показало, что чем больше был размер гиппокампа, тем в эффективнее данное животное обучалось пассивному избеганию.
Коэффициенты корреляции достоверно свидетельствовали о том, что размер гиппокампа (а возможно, какого-то из его отделов) определяет особенности выполнения выученного навыка (т.е. обучения как такового). Очень важно, что такая корреляция была получена в эксперименте без применения инвазивных методов, т.е. без прямого нарушения целостности мозга. Кроме того, поскольку исследуемая популяция мышей была генетически высокогетерогенной, можно было предположить, что обнаруженная скореллированная изменчивость обоих признаков (размер гиппокампа и эффективность научения) по крайней мере частично имела генетическую основу.
Морфометрические исследования (т.е. количественная оценка общих размеров) ряда отделов гиппокампа у мышей и крыс разных генотипов подтвердили существование достоверных межлинейных различий. Первоначально для анализа была выбрана условная реакция избегания в челночной камере (см. 3.2.2). Индивидуальную изменчивость темпов обучения этой реакции исследователи рассматривали как зависимую переменную (см.: Li рр etal., 1989; Schwegler, Lipp, 1995). В качестве независимой взяли вариабельность зоны окончания мшис-
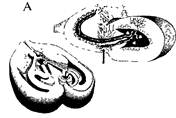
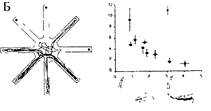
Рис. 9.2. Роль размера проекционной зоны iipMF гиппокампа мышей в формировании пространственного навыка поиска пищи в радиальном лабиринте.
А — схема строения гиппокампа; толстой стрелкой показана зона окончания iipMF; Б — слева: схема последовательных посещений мышью лучей радиального лабиринта, содержащих приманку, с небольшим числом повторных, ошибочных заходов; справа: график, отражающий зависимость между числом ошибочных заходов на 5-й день теста у мышей ряда инбредных линий, различающихся (нижняя схема) по относительному размеру проекции iipMF, который отложен по оси абсцисс графика (по: Lipp, Wolfer. 1995).
тых волокон, аксонов гранулярных клеток зубчатой фасции гиппокампа на базальных дендритах пирамидных нейронов поля САЗ (рис. 9.2А). Мшистые волокна оканчиваются в пирамидном слое поля САЗ крупными синаптическими бляшками. Зоны их окончаний формируют два четких синаптических поля, т.е. две области проекции. Одна из них располагается непосредственно над. пирамидными нейронами поля САЗ и называется супрапирамидным слоем. Вторая, меньшая по объему, располагается ниже или внутри слоя пирамидных клеток. Эта область называется слоем интра- и инфрапирамидных мшистых волокон, iipMF. Избирательная окраска именно этой структуры (метод Тимма) позво-пяет с высокой точностью определить ее размеры. У крыс Римских линий (см. выше), резко различающихся по скорости формирования навыка избегания удара тока, была выявлена отрицательная корреляция между площадью проекции мшистых волокон iipMF и способностью к обучению этой реакции. У мышей ряда инбредных линий, в том числе линий DBA/2J, СЗН/Не (размеры их зон проекций мшистых волокон схематически представлены на рис. 9.2Б справа), также была обнаружена сильная и высокодостоверная отрицательная (-0,92) корреляция площади UpMFn показателей обучения в челночной камере. У гибридов второго поколения от скрещивания этих линий корреляция может сохраниться только в случае, если ассоциация обоих признаков неслучайна. В эксперименте индивидуальная корреляция способности к обучению и площади iipMF у гибридов оказалась высокой.
Е На большом и разнообразном экспериментальном материале | было показано, что крысы и мыши тем лучше обучаются данному в навыку, чем меньше у них площадь iipMF.
Напомним, что выработка навыка избегания наказания в челночной камере — это типично лабораторный тест, аналога которому в естественном поведении грызунов практически нет. Его отрицательная корреляция с размером определенного отдела мозга еще ничего не говорит о функциональной значимости этой структуры. В этом отношении значительно больший интерес представляло исследование таких же корреляций в тестах, более адекватных экологической специализации крыс и мышей.
Для выяснения участия генотипа в формировании когнитивных способностей животных более информативными оказались данные о корреляции размера iipMF с успешностью формирования навыка обучения в радиальном лабиринте (см. 3.4.2.1), которое требует формирования пространственных представлений (формирования «мысленного плана» лабиринта, см. 3.4). В таких экспериментах была обнаружена достоверная положительная корреляция размера iipMF и обучаемости мышей пространственному навыку (Schwegler, Lipp, 1995). На рис. 9.2Б слева показана схема перемещения мыши по радиальному лабиринту при решении задачи, а также график зависимости успешности выполнения навыка от размеров данной зоны синаптических окончаний. Под графиком схематически изображены размеры проекций мшистых волокон у мышей двух линий (см. выше). Выполнение теста Морриса (обучение в водном лабиринте, см. 3.4.2.2), точнее, «прочность» сформированной пространственной памяти, положительно коррелирует с размером iipMF (Schwegler, Lipp, 1995).
g Тесты на способность к обучению на основе формирования представления о пространстве и о своем положении в нем животные усваивают тем успешнее, чем больше у них размер проекции мшистых волокон гранулярных клеток на базальныхдендритах пирамидных нейронов поля САЗ гиппокампа.
Были проанализированы корреляции успешности выполнения «пространственных» и непространственных тестов с размерами и других областей гиппокампа (не только с iipMF), однако таких фенотипических корреляций с толщиной отдельных слоев гиппокампа в его разных участках, т.е. с числом клеточных элементов и мощностью дендритных стволов пирамидных клеток, не обнаружено (Schwegler, Lipp, 1995).
|] Итак, обнаруженные нейроморфологические и поведенческие корреляции дают основание утверждать, что данная область синаптических окончаний (соединяющая гиппокамп с областью эн-торинальной коры и с новой корой) играет принципиально важ-ную, ключевую роль в осуществлении и/или модуляции процес-
li сов обучения разных типов.
Эти результаты были получены благодаря широкому использованию в лабораторных тестах генетически охарактеризованных живот-
ных, а также применению основных методов анализа генетических различий. В настоящее время исследование когнитивных способностей животных в этом тесте является одним из ведущих подходов в оценке особенностей поведения трансгенных животных и мышей-нокаутов. Детальнее с этими вопросами можно познакомиться в работе Lipp, Wolfer (1998), а также в материалах симпозиума «Behavioral Phenotyping of Mouse Mutants» (Cologne, 2000).
Использование трансгенных мышей при исследовании роли генотипа в процессах обучения и памяти. Методы генной инженерии и молекулярной биологии сделали возможным получение так называемых трансгенных животных. Как говорилось выше, такие исследования — это реализация подхода «от гена к поведению». Как правило, это физиологический и биохимический анализ, а также исследование поведения искусственных мутантов, у которых был видоизменен определенный участок генома. Для их создания в геном животного (в настоящее время используются почти исключительно мыши) вводится новый генетический материал. Таким материалом может быть или участок ДНК, кодирующий измененный ген, уже имеющийся у реципиента, ген от жи-ютного другого вида (например, крысы), либо генетическая конструкция, которая выключает какой-либо из генов реципиента (см.: Льюин, 1987; Jones, Mormede, 1999).
Выделенный фрагмент ДН К вводится в геном на ранней стадии эмбрионального развития. С методами введения можно ознакомиться в специальных руководствах. В результате соответствующих манипуляций формируются так называемые химерные животные. Нередко оказывается, что гомозиготные по новой мутации особи (мыши с обоими мутантными аллеями) нежизнеспособны, и ее удается поддерживать только в гетерозиготном состоянии. Однако чаще всего популяция трансгенных мышей представляет собой смесь из животных дикого типа (гомозиготных по нормальному аллелю гена), гетерози-готных особей (имеющих один нормальный и один мутантный аллель) и гомозиготных по мутантному аллелю. Генотип каждого животного можно определить методом полимеразной цепной реакции или иным методом, подвергнув анализу небольшой кусочек ткани животного (обычно для этого отрезают кончик хвоста). Иногда гомозиготные носители нового гена внешне отличаются от нормальных собратьев.
Специальные молекулярно-биологические приемы должны обеспечить достаточно надежную экспрессию новой ДНК в геноме реципиента. В противном случае введенный в геном фрагмент может сохраняться в латентном виде, не обнаруживая себя.
Наиболее часто эксперименты по получению искусственных мутантов или животных-нокаутов проводятся с целью выяснить роль в организме того или иного белка, чаще всего обладающего ферментативной активностью, или белков-рецепторов клеточной поверхности. При работе с нейрогенами, т.е. с генами, которые экспрессируются («работают») в мозге, наибольшее число исследований проведено с выключением белков-рецепторов, избирательно связывающихся с нейромедиа-торами и другими молекулами, влияющими на режим синаптической
передачи в нейронах разных структур мозга. В настоящее время насчитывается много сотен мышей-нокаутов с инактивированными генами разных функциональных групп, и их число продолжает расти.
Рассмотрим результаты наиболее известных исследований, в которых оценивали влияние выключения нейрогенов на процессы обучения и памяти.
Экспериментальные схемы обучения, которые используются для тестирования запоминания у лабораторных мышей и крыс, позволяют с большой надежностью проанализировать влияние какого-либо фактора на краткосрочную или долгосрочную память и на процесс собственно усвоения навыка. В качестве «навыка» обычно выбирают выполнение животным простой двигательной реакции или, наоборот, невыполнение (торможение) такой реакции. В целях большей четкости эксперимент строят таким образом, чтобы усвоение навыка происходило при единственном сочетании условного и безусловного раздражителей. Достаточно популярны в таких исследованиях оценка синаптической проводимости гиппокампа электрофизиологическими методами и формирования пространственного навыка в тесте Морриса.
Долговременная посттетаническая потенциация. Важным модельным объектом для изучения процесса обучения стала так называемая долговременная посттетаническая потенциация (long term potentiation, LTP).
LTP — это одно из проявлений синаптической пластичности (т.е. изменения проводимости синапсов), происходящее в результате длительной бомбардировки слоя пирамидных нейронов гиппокампа (область СА1) электрическим раздражением аксонов, оканчивающихся на этих нейронах (т.е. искусственным путем).
Подобные эксперименты в большинстве случаев проводятся на так называемых переживающих срезах гиппокампа, помещенных в специальную питательную среду, т.е. in vitro. В результате потенциации клетки гиппокампа начинают активно реагировать на раздражения, неэффективные до нанесения электрической стимуляции. Для этого процесса (как и для разных форм обучения) необходим целый ряд условий:
• активация так называемых NMDA-рецепторов (т.е. белковых молекул, расположенных в мембране нейронов и изменяющих ее проводимость при соединении с М-метил-О-аспартатом, ~ веществом, имитирующим эффект нейромедиатора);
• повышение внутриклеточного уровня ионов кальция (Са^), что в свою очередь обеспечивается определенным каскадом событий с участием системы так называемых вторичных посредников (циклического аденозинмонофосфата — С-АМР и др.);
• участие ряда ключевых ферментов (протеинкиназа- С, Сй^-калмо-дулинзависимая протеинкиназа II — СаМКИн тирозинкиназа).
Течение LTP в гиппокампе мышей-нокаутов с выключенными генами, кодирующими такие белки, сильно видоизменялось, однако она полностью не исчезала. Одновременно у этих мышей было нарушено формирование пространственного навыка в тесте g Морриса.
Гистологическое исследование гиппокампа показало у них нарушения в расположении гранулярных клеток зубчатой фасции, т.е. тех нейронов, аксоны которых образуют iipMF.
I) Мыши с искусственной мутацией гена Со MKII нормально обучались навыку отыскания безопасного убежища при наличии сигнальных раздражителей, но не могли усвоить этот навык, когда для этого требовалось формирование пространственных представ-
В лений, т.е. «пространственной карты» (см. 3.4).
К тому же у них при ритмическом электрическом раздражении гиппокампа с частотой 5—10 в сек (т.е. с частотой тета-ритма, как правило, присутствующего в суммарной электрической активности гиппокампа при исследовательском поведении) LTP не наступала, в то время как при высокочастотном раздражении она развивалась нормально. В норме у мышей могут развиваться обе формы LTP.
Можно проанализировать, как сказывается на процессе обучения противоположное генетическое изменение — «сверхэкспрессия» какого-либо гена. Специальными приемами можно усилить работу гена не во всем мозге, а только в определенных его участках. Такие вполне жизнеспособные животные демонстрируют поразительные видоизменения поведения (Mayford et al., 1995).
ез Сверхпродукция белка NMDA-рецептора типа 2В в переднем
Е мозге мутантных мышей сопровождалась резким усилением спо-р собности к обучению в ряде тестов (так же как и усилением LTP).
Мутантные мыши превосходили контрольных по усвоению навыка «застывания» (freezing response) при тестировании после однократного применения удара электрического тока, и у них было более прочное запоминание этой условной реакции. Обучение в тесте Морриса у этих мышей также было более эффективным. Данные наблюдения позволяют сделать вывод, важный для нейрофизиологии обучения:
В изменение эффективности NMDA-проводимости (и видимо, изменение интенсивности последующих процессов в нейроне) одинаково влияет на формирование разных навыков и, следовательно, представляет собой одно из универсальных звеньев в про-
С цессе обучения.
Известно, что формирование памяти — это ступенчатый процесс.
На животных разного филогенетического уровня показано, что в этом
процессе выделяются по меньшей мере две четкие стадии:
* краткосрочная память, которая не страдает от введения веществ подавляющих синтез белка или образование молекул РНК;
* долгосрочная память — ее формирование может быть блокировано введением этих веществ.
Переход от краткосрочной памяти к долгосрочной — консолидация — сопровождается активацией генетического аппарата или экспрессией новых генов, которые до этого были неактивны (репрессированы).
Считается установленным, что долговременная память связана с изменениями в структуре синоптических белков. Такие изменения осуществляются в результате целого каскада событий, принципиальным моментом которых является фосфорилирование, т.е. присоединение радикала неорганического фосфата к целому ряду белков. Этот биохимический процесс напрямую связан с изменениями в синапсах при их активации, он универсален и, как говорилось выше, принципиально сходен у животных разного уровня эволюционного развития. Выключение из каскада одного из его звеньев (путем «нокаута» соответствующего гена или, наоборот, усиления его работы) дает возможность оценить изменения в процессах собственно «следа» памяти.
Одним из наиболее известных примеров таких изменений у мышей-нокаутов является выключение гена, кодирующего белок CREB (Bourchaladze et al., 1994). CREB (c-AMP response element binding protein) относится к так называемым факторам транскрипции, или регулятор-ным белкам.
Е Мыши-нокауты по гену CREB достаточно эффективно обучались и хорошо запоминали навык в интервалах «работы» краткосрочной памяти (30 и 60 мин после сеанса обучения). Если же сохранение навыка тестировали в сроки, когда должна «работать» долгосрочная память (через 2 ч), то его воспроизведение было сильно нарушено.
Кроме этого, долговременная потенциация (LTP) у мышей с отсутствием гена, кодирующего белок CREB, развивалась аномально в тех же временных пределах. Через 2 ч после воздействия, вызывающего LTP, в срезах гиппокампа таких животных все ее проявления уже отсутствуют, тогда как в срезах мозга нормальных мышей сохраняются.
Совокупность данных, полученных на животных разного уровня развития, позволяет в настоящее время считать, что экспрессия транскрипционного фактора CREB, который активирует гены, прямо связанные с формированием памяти, и ряда других генетических элементов является важным этапом записи следа памяти в мозге.
Date: 2015-05-19; view: 690; Нарушение авторских прав