Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Роль генотипа в формировании способности к рассудочной деятельности
|
|
При тестировании элементарной рассудочной деятельности были получены многочисленные свидетельства вариативности (изменчивости) уровня выполнения этого теста среди животных одного вида. Л. В. Крушинский и его сотрудники в 60-70-е годы XX века проанализировали способность животных многих видов к экстраполяции направления движения стимула, т.е. их умение оперировать закономерностями перемещения предметов (см. гл. 4).
Сравнительные исследования поведения животных разных видов позволили сделать заключение, что уровень рассудочной деятельности тем выше, чем сложнее мозг животного (см. гл. 8). Однако для изучения физиолого-генетических основ этого феномена было необходимо исследовать животных одного вида, и наиболее подходящими объектами такой работы казались лабораторные грызуны, хорошо изученные как в физиологических, так и в генетических аспектах. Но именно у грызунов способность к экстраполяции оказалась развита слабо, в частности у лабораторных крыс и мышей она обнаруживалась далеко не всегда.
Экспериментальные данные о существовании генетических различий в способности животных к решению элементарных логических задач были получены в лаборатории Л. В. Крушинского при сравнении способности к экстраполяции у диких и доместицированных (одомашненных) форм лисицы и серой крысы. Дикие «красные» лисицы отличались высоким уровнем правильных решений теста на экстраполяцию. В то же время одомашненные черно-серебристые лисицы, в том числе и мутантные по цвету шерсти, разводившиеся в неволе в течение многих десятков поколений, выполняли этот тест с достоверно более низкими показателями, чем их дикие сородичи.
Рисунок 9.1А показывает успешность решения данного теста лисицами обеих групп. Доля правильных решений (на рисунке — высота столбиков) была выше у диких (1) лисиц, по сравнению с одомашненными (2—5). Очень высокий уровень правильных решений теста на экстраполяцию (даже при его первом предъявлении) наблюдали у прирученных диких крыс-пасюков, хотя эти показатели быстро снижались уже в течение первого опытного дня (т.е. при 6—8 предъявлениях теста). Лабораторные же крысы (линии Крушинского—Молодкиной
/KM), Wag, August и их гибриды между собой) оказались вообще неспособными к решению задачи на экстраполяцию. Доля правильных решений у них не превышала 50%-го уровня, т.е. они выбирали направление обхода ширмы чисто случайно, не руководствуясь информацией о направлении перемещения корма. В то же время гибриды первого поколения от скрещивания диких крыс с лабораторными обнаружили высокий уровень решения этой задачи, достоверно превышающий случайный уровень (Крушинский, 1986). Эти соотношения можно видеть на рис. 9.1 Б, где 1 и 2 — показатели диких крыс и их гибридов, 3-6 — соответственно крысы линий KM, WAG, Aug и гибридов KM x Aug.
И лабораторные крысы, и черно-серебристые лисицы, хотя и ведут свое происхождение от соответствующих диких форм, в течение многих поколений разведения в неволе не испытывали действия естественного отбора. Иными словами, в популяциях таких животных не было «выживания наиболее приспособленных», и соответственно доля животных, способных к быстрым адекватным реакциям на меняющиеся внешние условия, оказалась уменьшенной. Отражением этого можно считать снижение доли правильных решений теста на элементарную рассудочную деятельность.
Л. В. Крушинский (1986) предполагал, что в случае прекращения действия естественного отбора при размножении животных в неволе разрушаются сложные полигенные системы (или «коадаптированные комплексы»), которые в естественных условиях обеспечивают приспособление животных (через механизмы поведения) к изменяющимся и часто неблагоприятным внешним условиям.
Среди лабораторных мышей также были обнаружены генетические группы, у которых доля правильных решений задачи достоверно превышала случайную. Это были мыши с робертсоновской транслокацией (слиянием) хромосом Rb(8,l7)Jfern. В начале этого исследования способность к экстраполяции была проанализирована у значительного числа мышей с различными нарушениями кариотипа, в частности с робертсоновскими транслокациями разных хромосом. У животных с большинством таких мутаций доля правильных решений теста также не отличалась от 50%-го случайного уровня. В то же время мыши, у которых было слияние хромосом 8 и 17 (транслокации Rb(8,17)llem и Rb(8,17)6Sic; оно возникло совершенно независимо и найденно в разных лабораториях), оказались способными к экстраполяции. Рис. 9.1В показывает, что мыши со слиянием хромосом 8 и 17 (3-5) достоверно решали задачу на экстраполяцию, тогда как мыши с нормальным кариотипом (1,2) и со слиянием других хромосом (6) задачи не решали.
Мыши с этой хромосомной мутацией, в течение более 20 лет разводившиеся в нашей лаборатории, устойчиво показывали отличный от случайного уровень решения задачи на экстраполяцию. Позднее мы исследовали этот вопрос с использованием уникальной генетической
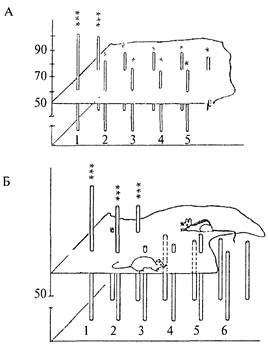
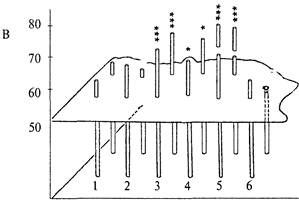
Рис. 9.1. Успешность решения теста на экстраполяцию животными разных генетических групп (пояснения в тексте).
А — решение теста лисицами; Б — крысами; В — мышами. Высота столбца соответствует доле правильных решений задачи при первом (ближний ряд) и многократных (дальний ряд) предъявлениях задачи.
модели — мышей 4 инбредных линий, которые попарно различались либо по генотипу (СВА и C57BL/6J), либо по наличию или отсутствию этой транслокации (Полетаева, 1998).
р Эксперименты с животными этих линий показали, что усиление способности к решению теста на экстраполяцию и другие особенности поведения, а также особенности обмена катехола-минов у этих мышей связаны именно с наличием в их кариотипе С данной робертсоновской транслокации.
Возможно, что причиной, лежащей в основе этих изменений в функции ЦНС при данной хромосомной перестройке, могут быть изменения в пространственном расположении генетического материала в интерфазном ядре, возникшие как следствие слияния хромосом.
Данные по различиям способности к экстраполяции у животных, отличающихся друг от друга генетически, естественно, не стоят особняком, а являются частью огромной «базы данных», созданной к сегодняшнему дню учеными, работающими в области генетики поведения.
Генетические исследования затрагивают практически все формы поведения, в том числе и способность к обучению, и способность к формированию пространственных представлений. Для того чтобы вкратце познакомиться с этим материалом, необходимо сначала дать краткий очерк основных методологических особенностей данного направления. Далее приводятся примеры использования генетических методов для изучения когнитивных способностей животных, а также краткое описание исследований генетических закономерностей психических способностей человека.
Date: 2015-05-19; view: 512; Нарушение авторских прав