Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Субмембранная система клетки
|
|
Субмембранная система клетки – специализированная периферическая часть цитоплазмы (в животных клетках она называется эктоплазма). Здесь практически отсутствуют органоиды. В эктоплазме сосредоточены ферментные системы трансмембранного транспорта, гликолиза. Эктоплазма обладает повышенной вязкостью. В эктоплазме располагается опорно-сократимая система.
Опорные (скелетные) фибриллярные структуры представлены промежуточными филаментами толщиной около 10 нм. Промежуточные филаменты образованы разнообразными белками: прекератин, виментин, десмин и другие.
Сократимые структуры образованы микрофиламентами (в их состав входят: актин, миозин, a–миозин, тропомиозин) и микротрубочками (которые на 80 % состоят из a– и b–тубулина, кроме того обнаруживаются динеин и другие белки). Соотношение между микрофиламентами и микротрубочками непостоянно: например, в микроворсинках клеток кишечного эпителия преобладают актиновые микрофиламенты, а в субмембранных структурах клеток ресничного эпителия преобладают тубулин–динеиновые микротрубочки. У инфузорий в равной степени представлены и микрофиламенты, и микротрубочки.
В специализированных клетках элементы опорно-сократительного аппарата носят исторически сложившиеся названия, например, в нейронах выделяют нейротрубочки и нейрофиламенты, а периферический слой цитоплазмы называется аксоплазма.
2. Рецепторы мембран
В состав плазмалеммы входят разнообразные молекулярные комплексы. Часть из них проявляет свойства рецепторов – структур, способных воспринимать изменения во внешней среде. Различают две группы рецепторов: неспецифические и специфические.
Неспецифические рецепторы способны связываться с самыми разнообразными веществами и частицами, например, бактериями, вирусами, белками. К ним относятся рецепторы на поверхности макрофагов, на поверхности мембран одноклеточных организмов.
Однако наибольший интерес представляют специфические рецепторы, способные реагировать только на строго определенные факторы. Рассмотрим действие специфических рецепторов на примере иммунной системы позвоночных животных, в основе которой лежит взаимодействиях антигенов и антител.
Антигены – разнообразные вещества, способные связываться с определенными белками – антителами. Например, антигены, определяющие группы крови у человека, могут быть и гликолипидами, и гликопротеинами. Антигены имеют специфические участки (антигенные детерминанты), которые и определяют их антигенные свойства. Каждый антиген регулирует синтез соответствующего (комплементарного) антитела.
Антитела (иммуноглобулины) вырабатываются специализированными клетками – плазмоцитами, которые образуются из В-лимфоцитов. Для плазмоцитов характерна хорошо развитая эндоплазматическая сеть, у них высокий уровень обмена веществ, повышенное содержание полисом – это настоящие фабрики антител. Время их жизни не превышает нескольких суток.
Антитела представляют собой гликопротеины со сложной четвертичной структурой. Молекулярная масса изменяется от 160 тысяч до 900 тысяч D (а.е.м.).
Структурно-функциональными единицами антител являются мономеры, состоящие из двух длинных (тяжелых – H) и двух коротких (легких – L) полипептидных цепей, связанных между собой дисульфидными связями. «Карманы» между тяжелыми и легкими цепями являются активными центрами, непосредственно образующими связи с антигенами. Активный участок антитела носит название Fab –участка. Основная часть антитела способна встраиваться в мембраны клеток и носит название Fc –участка.(Рис. 54).
У человека выделяют 5 классов иммуноглобулинов, различающихся по структуре и функциям: G, M, A, E, D. Молекулы иммуноглобулинов G, Е, D представлены мономерами, молекула иммуноглобулина М состоит из 5 мономеров, а молекула иммуноглобулина А может состоять из одного или двух мономеров. Антитела как сложные органические вещества также являются антигенами, т.е. могут вызывать синтез комплементарных им антител.
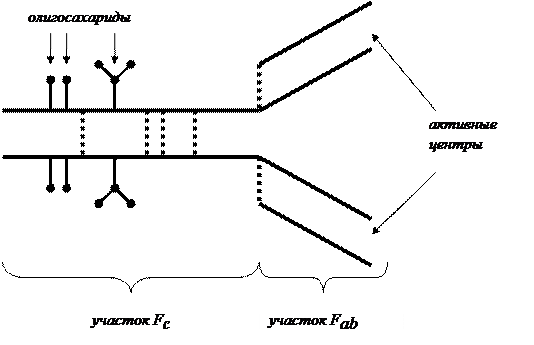 |
Рис. 54. Структурно-функциональными единицами антител.
Антитела делятся на нормальные и иммунные. Например, у людей с 0 (I) группой крови, у которых отсутствуют антигены А и В, вырабатываются нормальные иммуноглобулины (агглютинины) a и b, которые относятся к классу IgM. Эти антитела не проходят через плаценту. При иммунизации таких людей антигенами А и В у них вырабатываются иммунные иммуноглобулины (изоантитела) a и b, которые относятся к классу IgA. Эти антитела проходят через плаценту и могут вызывать иммунологический конфликт между организмом матери и плодом.
Гуморальный иммунитет и антитела в филогенезе впервые появляются у рыб. И только у плацентарных млекопитающих появляются все 5 классов антител. Однако клеточный иммунитет имеется и у высших беспозвоночных. У насекомых существует клетки–гематоциты, которые с помощью рецепторов распознают чужеродные антигены и блокируют их. С помощью этого механизма происходит изоляция личинок паразитов в теле насекомых.
Структура антител очень изменчива. Поэтому существует несколько уровней их разнообразия: изотипы, аллотипа, идиотипы, вариотипы.
Изотип – группа иммуноглобулинов данного класса, характерная для определенного вида организмов. Например, иммуноглобулин G кролика и иммуноглобулин G человека – разные изотипы иммуноглобулина G. Соответственно, иммуноглобулин G кролика является антигеном для человека и наоборот. Изотип определяется особенностями участка Fc.
Аллотип – группа иммуноглобулина данного класса, характерная для определенной особи. Например, иммуноглобулин G у Иванова и иммуноглобулин G у Петрова – разные аллотипы иммуноглобулина G человека. Аллотип определяется генотипом особи.
Идиотип – конкретные одинаковые молекулы иммуноглобулина данного класса, продуцируемые одним клоном клеток. В ходе дифференциации В–лимфоцитов каждая клетка приобретает способность синтезировать лишь один идиотип. Особенности идиотипа определяются строением вариабельных участков легкой и тяжелой цепей.
Вариотип – группа иммуноглобулинов данного класса, отличающаяся от подобных групп последовательностью аминокислот, которая обычно консервативна (инвариантная последовательность).
Таким образом, антитела обеспечивают образование самых разнообразных комплексов [антиген-антитело].
Антитела могут находиться в сыворотке крови (сывороточные антитела), но могут связываться своим Fc –участком с мембранами лимфоцитов. Такое связанное с мембраной антитело является рецептором антигена.
В то же время на поверхности многих клеток имеются и рецепторы для Fc –участка антитела, Fc –рецепторы – гликолипопротеины или гликопротеины разнообразной структуры, встроенные в мембрану самых разнообразных клеток.
Антигенными свойствами обладает не только гликокаликс, но и поверхность клеточных стенок, а также поверхность оболочек вирусов. У некоторых вирусов (герпеса, гриппа, ВИЧ и др.) образуется дополнительная оболочка из плазмалеммы клеток хозяина, что снижает антигенные свойства этих вирусов.
При образовании комплексов [антиген-антитело] или [рецептор-белок] на поверхности клеток изменяется поверхностное натяжение плазмалеммы, что приводит к изменению физиологического состояния клетки.
3. Транспорт веществ через мембраны
Транспорт веществ через мембраны может осуществляться по градиенту концентрации (от большей концентрации к меньшей) и против градиента концентрации (от меньшей концентрации к большей).
Вода, многие липиды, ионы магния, хлора и некоторые другие вещества свободно перемещаются через мембраны по градиенту концентрации путем простой диффузии (без затраты энергии).
Углеводы, некоторые аминокислоты, некоторые липиды и некоторые ионы перемещаются по градиенту концентрации путем облегчённой диффузии (с помощью белков–переносчиков с предварительной затратой энергии).
Ионы водорода, калия, натрия, кальция и некоторые органические вещества перемещаются через мембраны против градиента концентрации (с помощью белков–переносчиков и с затратой энергии). Такой транспорт веществ называется активным.
Крупные молекулы и частицы поглощаются клеткой путем фагоцитоза, а капли раствора – путем пиноцитоза. Фаго- и пиноцитоз объединяются под общим названием эндоцитоз. Эндоцитоз характерен для специализированных клеток амебоидного типа. Эндоцитозу предшествует фиксация молекул и частиц на внешней поверхности мембраны с помощью специфических и неспецифических рецепторов.
Транспорт крупных молекул и частиц из клетки наружу называется экзоцитоз. При экзоцитозе образуются экзоцитозные пузырьки из внутриклеточных мембран, которые, приближаясь к плазмалемме, изменяют ее поверхностное натяжение.
Пример активного транспорта: натрий–калиевый насос
Плазмалемма пронизана сложным белком–переносчиком (K–Na–АТФаза). В исходном состоянии этот переносчик может присоединять с внутренней стороны мембраны три иона Na+. Эти ионы являются аллостерическим активатором и изменяют конформацию активного центра АТФазы. После такой активации АТФаза способна гидролизовать одну молекулу АТФ, причем, фосфат-ион фиксируется на поверхности переносчика с внутренней стороны мембраны. Выделившаяся энергия расходуется на новое изменение конформации АТФазы, после чего три иона Na+ и ион PO43- оказываются на внешней стороне мембраны. Здесь ионы Na+ отщепляются, а PO43- замещается на два иона К+. После этого конформация переносчика изменяется на первоначальную, и ионы К+ оказываются на внутренней стороне мембраны. Здесь ионы К+ отщепляются, и переносчик вновь готов к работе. В итоге на внешней стороне мембраны накапливается избыточный положительный заряд.
Пример облегчённой диффузии: транспорт глюкозы в клетку
Энергия избыточного положительного заряда поверхности мембраны используется в работе натрий–зависимых переносчиков глюкозы, аминокислот и некоторых ионов.
Ионы Na+ присоединяются к внешней части переносчика и активируют его. Затем к внешней части переносчика присоединяется молекула другого вещества (в частности, глюкозы), что приводит к изменению конформации переносчика. В результате ионы Na+ вместе с глюкозой оказываются на внутренней стороне мембраны, где происходит их отщепление. Переносчик изменяет свою конформацию на исходную и вновь приступает к работе. Активность натрий-зависимого переносчика глюкозы регулируется гормоном инсулином.
При работе K–Na–АТФазы и натрий–зависимого переносчика глюкозы на внутренней стороне плазмалеммы накапливается избыток ионов K+, а на внешней стороне – избыток РО43-. Однако эти ионы способны к простой диффузии через мембраны, что приводит к постепенному выравниванию концентраций.
Пример эндо- и экзоцитоза: транспорт холестерина
Холестерин играет важную роль в поддержании структуры мембран, на его основе синтезируются желчные кислоты и стероидные гормоны. В среднем около 1000 мг холестерина в сутки синтезируется у человека в клетках печени, а около 500 мг поступает с животной пищей. Избыток холестерина в организме приводит к изменению мембран, а в итоге – к развитию ишемической болезни.
В плазме крови холестерин (как и любой липид) находится в виде липопротеинового комплекса [холестерин–апопротеин]. На поверхности клеток печени имеются рецепторы этого комплекса. В результате образуется комплекс [холестерин–апопротеин–рецептор]. После образования такого комплекса изменяется поверхностное натяжение мембраны, и образуется впячивание – эндоцитозный пузырек. В итоге весь комплекс [холестерин–апопротеин–рецептор] оказывается внутри клетки.
Здесь комплекс разрушается: холестерин поступает в цитоплазму, белок–апопротеин гидролизуется до аминокислот, а рецепторы возвращаются на поверхность мембраны в составе экзоцитозного пузырька. Избыток холестерина в клетке хранится в виде его эфиров.
При избытке холестерина в клетках он подавляет свой собственный синтез – возникает отрицательная обратная связь: избыток холестерина в крови снижает синтез холестерина клетками печени.
Однако при значительном избытке холестерина в клетках подавляется синтез холестерин–апопротеиновых рецепторов. Тогда прекращается поступление холестерина из крови в клетку, и синтез холестерина клетками печени не тормозится – возникает положительная обратная связь: в крови сохраняется холестерин, полученный с пищей, а клетки печени продолжают синтез холестерина.
Некоторое время в плазме крови сохраняется избыток холестерина. При длительном насыщении организма холестерином его избыток не проникает в клетки печени, а откладывается на поверхности стенок кровеносных сосудов, что и приводит к развитию атеросклероза и ишемии.
При некоторых наследственных заболеваниях нарушается структура рецепторов (известно около 10 таких заболеваний). Тогда холестерин не может поступать в клетки и регулировать свой собственный обмен. В плазме крови накапливается избыток холестерина и за счет поступления с животной пищей, и за счет синтеза клетками печени. У гомозигот по таким заболеваниям ишемическая болезнь развивается еще в детстве, а у гетерозигот – примерно к 50 годам.
4. Межклеточные контакты
Межклеточные контакты условно делятся на собственно структурные контакты и функциональные взаимодействия.
Наличие толстых клеточных стенок часто затрудняет структурный контакт между клетками. Клеточные стенки у таких организмов пронизаны порами, через которые проходят тяжи цитоплазмы – плазмодесмы. Совокупность клеток, связанных плазмодесмами в единое целое, называется симпласт (цианобактерии, грибы и растения).
Ряд организмов на некоторых стадиях развития образует плазмодий – гигантскую многоядерную клетку. Плазмодий возникает в том случае, если ядра делятся (происходит кариокинез), а цитоплазма остается единой (отсутствует цитокинез). Стадия плазмодия встречается у многих простейших, слизевиков, низших грибов, а также у ряда прокариот.
В некоторых случаях многоядерные клетки образуются при слиянии одноядерных. В этом случае применяют название синцитий (синцитий хорошо выражен у некоторых губок и в покровах плоских червей).
Если же при объединении клеток границы между ними сохраняются, то такая структура называется псевдоплазмодий (например, у слизевиков).
Взаимодействия рецепторов мембран с антигенами и антителами являются основой функциональных контактов между клетками, которые обеспечивают жизнедеятельность клеток, тканей, органов и систем органов, а также поддержание гомеостаза, дифференцировку клеток многоклеточного организма у животных и т.д. Однако в некоторых случаях реакция организма на антиген может быть чрезмерной, например, при анафилаксии.
Анафилаксия, или гиперчувствительность – это немедленная реакция организма плацентарных млекопитающих на повторное воздействие сверхмалых доз антигенов. Симптомы анафилаксии у человека: одышка, частый пульс, падение давления, отеки... При анафилаксии происходит образование микротромбов в крови вблизи антигенов. Причиной анафилаксии могут быть самые разнообразные антигены, которые в данном случае называются аллергенами. При анафилаксии необходимо применять антигистаминные препараты или иные неспецифические средства.
При попадании любого вещества в организм оно захватывается макрофагами (А–клетками) с помощью неспецифических рецепторов (макрофаги сосредоточены в печени, в легочных капиллярах и в ряде других органов). В макрофагах поступившее вещество расщепляется на более простые структуры – происходит презентация антигена. Образовавшиеся антигены выводятся на поверхность макрофага и образуют комплекс [антиген–иммуноглобулиновый рецептор антигена] на поверхности В–лимфоцита. Данный В–лимфоцит должен обладать способностью продуцировать идиотип иммуноглобулина Е (IgE) именно к данному антигену, и именно этот идиотип является рецептором данного антигена.
Если антиген является белком, то в процессе образования комплекса на поверхности В–лимфоцита принимают участие Т–хелперы – определенный тип Т–лимфоцитов. (Именно Т–хелперы, или Т-4–лимфоциты поражаются вирусом СПИДа.)
После образования комплекса [антиген–рецептор] В–лимфоцит превращается в плазмоцит и начинается интенсивный синтез идиотипа IgЕ именно к данному антигену. Этот идиотип IgЕ попадает в плазму крови или в тканевую жидкость и присоединяется к своему рецептору Fc –участка на поверхности базофилов или тучных клеток. Комплекс [ Fc –рецептор IgЕ – идиотип IgЕ] очень устойчив и сохраняется в течение нескольких недель. Базофилы и тучные клетки содержат большое количество анафилактических медиаторов (гистамин, ацетилхолин и др.), но в обычных условиях эти медиаторы через мембрану не проходят.
При повторном попадании рассматриваемого антигена в организм вновь происходит его презентация макрофагами и последующая фиксация на поверхности базофилов или тучных клеток с помощью комплекса [ Fc –рецептор IgЕ – идиотип IgЕ]. Образуется комплекс [ Fc –рецептор IgЕ – идиотип IgЕ – антиген]. При этом снижается поверхностное натяжение мембраны, и медиаторы выбрасываются в плазму крови или тканевую жидкость.
Кроме анафилаксии у человека широко распространена другая группа аллергических заболеваний – разнообразные атопии. Атопии также связаны с усиленным продуцированием IgЕ, но обусловлены генетическими особенностями особи. При атопии комплексы идиотипов IgЕ на поверхности тучных клеток и базофилов образуются самопроизвольно, еще до первого контакта с антигеном.
Date: 2015-04-23; view: 2790; Нарушение авторских прав