Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Сознание и мозг
|
|
Новые функции, которые должен был принять на себя мозг человека в связи с развитием труда, отразились на изменении его строения. Коренное изменение характера деятельности — с переходом от жизнедеятельности к трудовой деятельности, всё более усложнявшийся характер этой деятельности и соответственно всё углублявшийся характер познания привели к тому, что над проекционными зонами, непосредственно связанными с периферическими сенсорными и моторными аппаратами, развились богатые ассоциативными волокнами зоны, служащие для более сложных синтезов. Сравнение мозга человека с мозгом обезьяны (см. рис. ниже) отчётливо выявляет эти сдвиги: у человека первичное зрительное поле, столь развитое у обезьян, заметно уменьшается, и вместе с тем значительно возрастают поля, с которыми связаны сложные синтезы зрительного восприятия (вторичное зрительное поле).
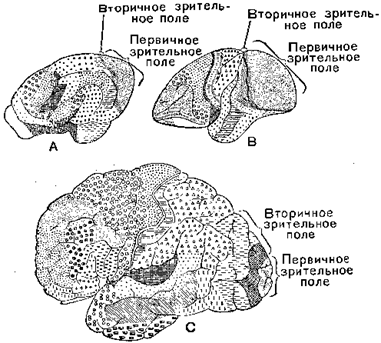
А — мозг медведя, В — оранга и С — человека
Согласно исследованиям Филимонова (Институт мозга в Москве), затылочная область коры занимает у оранга 21,5% всей поверхности большого мозга, а у человека всего 12%, что связано с более значительным развитием у человека новых полей, выполняющих более сложные функции.
Исследования Шевченко, посвящённые сравнительному изучению нижней париетальной области, являющейся носительницей особенно высоких функций, обнаружили, правда, — вопреки утверждению предыдущих авторов — наличие этих полей не только у антропоморфных, но и у церкопитеков, но вместе с тем они показали что у человека эта нижняя париетальная область сильно вырастает: равная всего 0,4% коры большого мозга у церкопитеков, 3,4% — у оранга и шимпанзе, она достигает 10% у человека.
С изменением строения и функции коры у человека связана и вырастающая роль коры — новый этап в процессе кортикализации. В то время как у всех позвоночных вплоть до хищных с мозговым стволом связаны ещё психические функции, у человека он является лишь рефлекторным аппаратом; психические процессы являются у него функцией коры — органа индивидуально приобретённых форм поведения.
В онтогенезе у человека головной мозг развивается из переднего отдела медулярной мозговой трубки, в которой постепенно образуются 5 мозговых пузырьков (телэнцефален, состоящий из двух полушарий, диэнцефален, или промежуточный мозг, мезэнцефален, или средний мозг, метэнцефален, или задний мозг, миэленцефален).
Полушария большого мозга образуются из конечного мозга (телэнцефалена), развивающегося из первичной рецепторной пластинки; из остальных отделов медулярной трубки развиваются друг над другом надстраивающиеся отделы мозгового ствола. Мозговой ствол включает в себя продолговатый мозг (образующийся из диэленцефалена), варолиев мост и мозжечок (из метэнцефалена), четверохолмие, ножки мозга (средний мозг), зрительные бугры (таламус, эпи- и гипоталамус) и коленчатые тела (межуточный мозг). Передние отделы мозгового ствола (зрительные бугры и коленчатые тела) совместно с узлами, расположенными в примыкающей к стволу нижней части полушарий большого мозга под корой, составляют так называемую подкорку.
Продолговатый мозг регулирует такие автоматические функции, как деятельность сердца, дыхание, перистальтику кишок, глотательные, а также рвотные движения, под воздействием поступающих к нему импульсов от органов кровообращения, дыхания, от языка, пищеварительного канала. Варолиев мост иннервирует слюноотделительные железы, мышцы лица и мышцы, поворачивающие кнаружи глазные яблоки. Мозжечок, функции которого ещё полностью не выяснены, играет во всяком случае существенную роль в управлении тонусом, в распределении напряжения скелетных мышц, необходимом для координации движений. Мозжечок — регулятор тонических сухожильных рефлексов. Координацию наших движений мозжечок осуществляет в сотрудничестве с вышележащими отделами нервной системы, в том числе и корой. В регулировании нашей моторики принимает участие и расположенное над мозжечком красное ядро, в которое поступают афферентные импульсы от разных органов чувств.
В четверохолмие, лежащее, так же как и красное ядро, в области среднего мозга, поступают афферентные волокна зрительного нерва (в переднее двухолмие) и невроны слухового нерва (в заднее двухолмие). Из четверохолмия исходит иннервация мышц, движущих глазное яблоко, аккомодационной мышцы и мышцы, суживающей отверстие зрачка.
Очень существенную роль играет подкорка, состоящая подобно спинному мозгу из сенсорной, моторной и вегетативной зон. Сенсорная зона — это зрительный бугор, куда сходятся афферентные пути от всех рецепторов тела. Всякие центростремительные импульсы — болевой, осязательный, вкусовой, обонятельный, слуховой, зрительный, прежде чем поступать в кору, попадают в зрительный бугор. Они могут переключаться здесь же в подкорке на её двигательные ядра (полосатое тело и бледный шар), в результате чего осуществляются движения непроизвольного автоматического или инстинктивного характера или направляются далее к коре полушарий головного мозга. Подбугровая область подкорки состоит из центров вегетативной нервной системы, которые изменяют состояние всех органов тела при возбуждении тех или иных органов чувств или коры полушарий. Управляя реакциями внутренних органов и в первую очередь сердца, подкорковые бугры играют существенную роль в аффективно-эмоциональных состояниях и при мышечной работе.
Кора представляет собой, как известно, как бы плащ или мантию, покрывающую большие полушария. Её сравнительно очень значительная поверхность (около 2000 кв. см)собрана в ряде складок, или борозд, и извилин (см. рисунки ниже). Из них основные: центральная, или роландова, борозда, расположенная приблизительно посредине полушарий; снизу от неё проходит сильвиева борозда, в лобной и височной частях проходят по три извилины — верхняя, средняя и нижняя лобные и височные извилины; в затылочной части выделяются первая, вторая и третья затылочные извилины, а в теменной две — верхняя и нижняя теменные извилины.
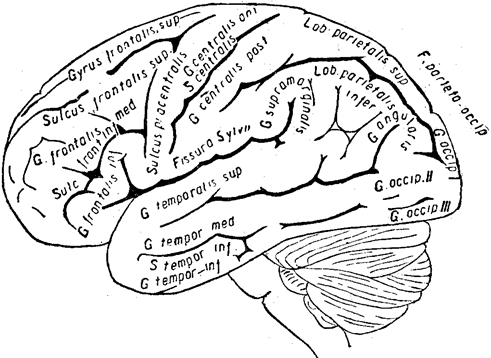
Наружная поверхность левого полушария головного мозга и мозжечка (по Рауберу)
Поскольку у человека органом сознательной деятельности является кора, вопрос о взаимоотношении психики и мозга сосредоточивается в первую очередь на вопросе о взаимоотношении психики и коры больших полушарий головного мозга.
Вопрос о взаимоотношении психики и коры выступает в науке конкретно, как вопрос о функциональной локализации, или локализации психических функций в коре.
Учение о локализации прошло через несколько фаз, смена которых ярко отражает борьбу двух течений, существующих и поныне в формах, соответствующих современному состоянию научного исследования. В начале XIX в. Галль, отражая психологическую теорию способностей, попытался построить френологию или органологию мозга. Он рассматривает мозг как совокупность мозговых органов, в которых он локализирует и такие сложные психологические образования, как чувство сравнения, память, родительская любовь, поэтический талант и т. д. В 40-х годах против френологии выступает Флуранс, который на основании своих опытов экстирпации частей мозга выдвигает то положение, что мозг является однородной массой, функционирующей как единый цельный орган. Основу современного учения о локализации кладёт открытие Брока, в 1861 г. обнаружившего, что разрушение третьей лобной извилины левого полушария связано с поражением речи.
Вслед за открытием Брока двигательного «центра» речи в ближайшие десятилетия последовал ряд других открытий. Вернике показал связь так называемой словесной глухоты с задней частью первой височной извилины,в которой был локализирован центр Вернике. Дежерин установил связь алексии (зрительной афазии) с gyrus angularis; Липман — связь апраксии (нарушение действия) с gyrus supramarginalis; Экснер — связь аграфии (нарушение письма) с задним концом второй лобной извилины. Далее была установлена связь зрительной агнозии (нарушение познания, узнавания) с поражением наружной поверхности затылочных долей,нарушения кожной чувствительности — с поражением задней центральной извилины и верхней теменной дольки. Путём экстирпации отдельных участков полушарий у собак и обезьян Мунк установил связь зрения с затылочными долями полушарий, слуха — с височными. Этот ряд исследований привёл к установлению ряда корковых рецепторных, сенсорных «центров».
С другой стороны, Гитциг и Фритч в 1871 г. показали, что раздражение электрическим током определённых участков коры сопровождается движениями определённых частей тела. Это привело к установлению эффекторных центров коры,расположенных с обеих сторон, впереди роландовой борозды. И рецепторные, и эффекторные центры являются проекционными центрами коры. Они связаны с периферической реакцией и эффектом. Таким образом, вырисовывалась как бы карта коры (см. рисунки ниже).

Центры на наружной поверхности головного мозга (по Раубер-Копшу). Штриховкой точками обозначены сенсорные центры, вертикальными чёрточками — двигательные центры

Центры на внутренней поверхности головного мозга (по Раубер-Копшу)
Для объяснения функционирования мозга, расчленённого таким образом на совокупность отдельных центров, нужно было дополнительно допустить существование фиксированных ассоциационных путей, или рефлекторных дуг, объединяющих эти центры. Это оказалось тем более необходимым, что, как показало исследование, нарушение речи, например, связано не только с поражением «центра» Брока, но и других центров в левом полушарии. Поэтому нужно было объяснить совместное действие нескольких центров в выполнении одной функции.
На этой основе установилась локализационная теория, господствовавшая в науке до последнего времени. Она сложилась в результате того, что над положительными фактическими данными исследования было воздвигнуто здание гипотез и теорий, отражающих те же методологические тенденции, которые господствовали и в тогдашней психологии. Представление о мозге, как совокупности или мозаике отдельных центров, соединённых между собой ассоциационными путями, отражало концепцию ассоциативной психологии, из которой по существу и исходила классическая локализационная теория. Представление же о том, что каждой психической функции, в том числе самым сложным, соответствует определённый центр, является своеобразной и наивной реализацией в физиологии головного мозга теории психофизического параллелизма.
Дальнейшую детализацию и уточнение учение о локализации получило благодаря исследованиям Флексига о миэлинизации (обложении мякотной оболочкой волокон) различных областей коры и исследованиям (К. Бродмана, Ц. и О. Фохтов, Экономо и Коскинаса) о миэло- и цитоархитектонике (о волоконцевом и клеточном составе) коры.
На основании миэлогенетических исследований, установивших различное время обкладывания мякотными оболочками волокон различных систем, Флексиг разделил всю кору на 36 гистологически отличных полей. Он при этом исходил из того положения, что функциональное формирование и функциональная зрелость центров связаны с миэлинизацией соответствующей системы волокон. Придя к тому заключению, что теменные и лобные доли, которые миэлинизируются наиболее поздно, лишены проекционных систем, соединяющих их с нижележащими центрами ствола, Флексиг сделал недостаточно обоснованный вывод о том, что эти поля являются ассоциационными центрами, в которых локализируются высшие психические функции. Дальнейшее изучение строения коры показало, что выделенные Флексигом ассоциационные сферы имеют не только сочетательные, но и проекционные системы и что отношения между различными областями коры значительно сложнее. Исследования, начало которым положили Мейнерт и Бетц, привели к новому учению об архитектонике коры.
Цитоархитектоника изучает клеточное строение коры, миэлоархитектоника — ход её волокон. Основной тип строения коры характеризуется наличием в ней 6 или (по О. Фохту) 7 клеточных и волоконцевых слоёв: 1) зональный (или молекулярный) слой, бедный клетками и состоящий главным образом из волокнистых сплетений; 2) наружный гранулярный, зернистый слой, изобилующий мелкими зернистыми, пирамидообразными клетками; 3) пирамидный слой, состоящий из средних по величине и относительно бОльших пирамидных клеток; 4) внутренний гранулярный слой с большим количеством мелких зернистых клеток; 5) ганглионарный слой, состоящий из крупных пирамидальных клеток, так называемых глубоких пирамид, достигающих особенного развития в передней центральной извилине, в которой Бетц обнаружил носящие его имя «гигантские» пирамидные клетки; 6) полиморфный слой, состоящий из клеток различной величины и формы; 7) веретённый слой (по О. Фохту), содержащий веретенообразные клетки и граничащий с белым веществом (см. рис.).
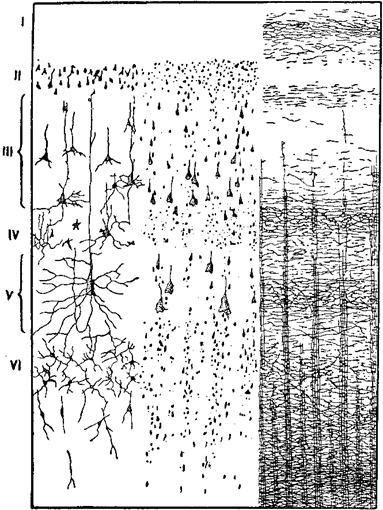
Слой нервных клеток и нервных пучков в коре большого мозга человека
Как показали исследования Ц. Фохт, цито- и миэлоархитектонические особенности коры в разных отделах её различны. На основании этих цито- и миэлоархитектонических особенностей различных частей коры К. Бродман составил карту коры, на которой он различает 52 корковых поля различной архитектоники (см. рисунки ниже). Дальнейшие исследования Ц. и О. Фохтов (1919 г.), Экономо и Коскинаса (1925 г.) привели к выделению в коре у человека до 200 корковых полей.

Ареальная карта по К. Бродману. Наружная поверхность

Ареальная карта по К. Бродману. Внутренняя поверхность
Изучение филогенеза мозга показало, что в филогенетическом ряду наблюдается всё возрастающая анатомическая дифференциация коры, причём всё большее развитие получают те участки, которые являются носителями особенно высоких функций.
Существенные результаты даёт и изучение онтогенетического развития архитектоники коры. Применённый впервые К. Бродманом принцип деления коры на основании изучения её онтогенетического развития (которое привело его к различению гомогенетической коры, приходящей в процессе своего онтогенетического развития к шестислойному строению, и гетерогенетической коры, не проходящей через стадию шестислойного расщепления) получил дальнейшее развитие у ряда советских учёных. Исследования И. Н. Филимонова, Г. И. Полякова, Н. А. Попова показали, что уже на ранних стадиях онтогенетического развития выступает деление коры большого мозга на три основные зоны: 1) изокортекс, 2) аллокортекс, включающий архикортекс и палеокортекс, и 3) определяющую алло- и изокортекс межуточную область. Наличие этого деления уже на ранних стадиях онтогенеза даёт основание заключить, что оно имеет существенное значение.
В результате современных исследований можно считать установленным, что кора состоит из гистологически различных, могущих быть индивидуально дифференцированными полей. Не должно было бы также подлежать сомнению и то, что с гистологическими особенностями связано и некоторое функциональное своеобразие. За это положение говорит, во-первых, накопившийся с открытием Брока ряд данных о связи нарушения определённых функций с поражением определённых участков коры. Исследования Павлова также показали, что разрушение у собаки различных участков коры влечёт за собой расстройство анализаторской деятельности различных сенсорных областей. Это положение подтвердилось и посредством других методов. Ц. и О. Фохты, раздражая электродом на мозгу обезьян различные участки, показали, что гистологически различные ими установленные поля дают различный моторный эффект. Аналогичные данные получил тем же методом, применённым им во время операций над мозгом человека, Ферстер, причём ему удалось констатировать таким способом и различия сенсорных полей.
Богатый материал для решения локализационной проблемы дают и клинические данные современной психоневрологии.
В итоге этого длинного ряда исследований было, таким образом, установлено, что мозг является не однородной массой, а гистологически и функционально качественно дифференцированным целым, в котором различные участки обладают определённым своеобразием, и что определённые функции оказываются в известной зависимости от гистологически определённых участков мозга, которые, следовательно, играют особенно существенную роль в их деятельности. Над этим бесспорным положением, опирающимся на факты, классическая локализационная теория воздвигла очень гипотетическое и шаткое построение, допустив, что каждая, даже сложная, психическая функция непосредственно продуцируется отдельным участком как её «центром».
Эта классическая локализационная теория в настоящее время основательно поколеблена исследованиями Х. Джексона, Г. Хэда, работами К. Монакова, К. Гольдштейна, К. Лешли и др. Оказалось прежде всего, что новые клинические данные о многообразных формах афазии, агнозии, апраксии не укладываются в классическую локализационную схему. С одной стороны, поражение так называемой речевой зоны в левом полушарии при более тщательном исследовании оказывается связанным с расстройством нетолько речи, но и других интеллектуальных функций. С другой стороны, нарушение речи, различные формы афазии связаны с поражением различных участков.
При оценке обоснованности выводов традиционной локализационной теории нужно, далее, учесть, что методом экстирпации на основе поражения различных участков коры могли быть установлены лишь «центры нарушения»,а не центры функционирования в собственном смысле. Из того факта, что поражение определённого поля влечёт за собой нарушение определённой функции, следует, что это поле играет существенную роль в выполнении данной функции; но это не означает, что оно является «центром», который сам продуцирует эту функцию, и что в ней не участвуют и другие поля. Надо поэтому различать между локализацией патологического симптома в определённом поле и локализацией функции в определённом «центре».
Это положение особенно заострил против классического учения о локализации К. Монаков. Как указал Монаков и подтвердил К. Лешли, значительные повреждения и других участков коры помимо тех, с поражением которых в первую очередь связано нарушение высших интеллектуальных функций, также влечёт за собой их нарушение, хотя и не столь глубокое. С другой стороны, и при разрушении основных для данной функции участков остальная часть коры по истечении некоторого времени до известной степени компенсирует дефект, принимая на себя замещающую роль.
Особенно существенное значение для локализационной проблемы имеет функциональная многозначность гистологически определённых полей коры. Даже разделению коры на отдельные сенсорные и моторные участки нельзя придавать абсолютного значения. Вся кора функционирует как чувствительно-двигательный аппарат, в котором имеется лишь местное преобладание одной из двух функциональных сторон — двигательной или сенсорной. Дифференциация функциональных особенностей связана с преобладающим развитием в данной области одного из двух основных слоёв коры (А. Якоб).
Опираясь на свои исследования о зрительной коре, а также лобной и центральных областях у крыс и обезьян, К. Лешли, выдвигающий положение о функциональной многозначности корковых полей, показал, что разрушение зрительной области коры ведёт к трём различным психологическим изменениям: 1) животное теряет способность различать формы; 2) наступает амнезия по отношению к прежде приобретённым навыкам; 3) приобретение новых сложных навыков оказывается тем труднее, чем обширнее повреждённая область. При этом Лешли установил, что причиной двух последних нарушений не является нарушение зрительных восприятий. Таким образом, зрительная кора обусловливает несколько различных функций, играя, однако, по отношению к ним различную роль. Признавая наличие в зрительной коре специального, строго локализированного механизма различения формы и менее строго локализированного механизма активирования подкорковых центров, Лешли считает, что свойство совместно со всеми другими участками коры осуществлять интеллектуальные функции является совершенно не специфическим: высшие интеллектуальные функции вовсе не локализируемы.
В отношении высших интеллектуальных функций ряд современных исследователей, как-то: К. Монаков, К. Гольдштейн, К. Лешли (в ранних своих работах, от которых сейчас он отошёл), в своей борьбе против традиционной локализационной теории делают своеобразный зигзаг в противоположном направлении — обратно к идеям М. Ж. П. Флуранса. Так, Монаков принимает локализацию только «входных» и «выходных ворот»,т. е. мест вхождения в кору рецепторных путей и выхода из неё эффекторных. Для всех высших, более сложных психических функций Монаков признаёт только «хроногенную» локализацию. Он, конечно, прав в том, что сложную интеллектуальную функцию нельзя локализировать в каком-нибудь одном центре, что в её осуществлении должны принять участие, каждый со своим вкладом, нервные элементы, распределённые на обширной территории мозга, действие которых объединяется в одной временнОй структуре. Правильной также в своей основной идее является тенденция К. Монакова локализировать каждую функцию во времени, путём отнесения её к определённой стадии в генетическом ряду. Но локализация только во времени, принципиально противопоставляемая всякой локализации в пространстве, ведёт в конечном счёте к идеалистическому отрыву психики от её материального субстрата.
К. Лешли в своих ранних работах ещё заострённее переходит от правильного положения о функциональной многозначности корковых полей к отрицанию какого бы то ни было «соответствия структурных и функциональных единиц». Он считает, что можно говорить лишь о количественном соответствии между объёмом мозгового повреждения и степенью расстройства интеллектуальной функции.
В результате друг другу противостоят две непримиримые и, пожалуй, равно неправомерные концепции: согласно одной, мозг представляет мозаику или механическую сумму разнородных центров, в каждом из которых локализирована особая функция; согласно другой, мозг функционирует как целое, но такое, в котором все части функционально равнозначны, так что, архитектонически чрезвычайно дифференцированный, он функционально представляется однородной массой.
Учёт всех фактических данных, экспериментально установленных как одной, так и другой стороной, допускает только одно решение: в сложных психических функциях у человека участвует значительная часть коры или вся кора, весь мозг как единое целое, но как целое и функционально и гистологически качественно дифференцированное, а не как однородная масса. Каждая его часть участвует в каждом целостном процессе более или менее специфическим образом. Для сложных интеллектуальных функций не существует «центров», которые бы их продуцировали, но в осуществлении каждой из них определённые участки мозга играют особенно существенную роль. Для интеллектуальной деятельности особенно существенное значение имеют, очевидно, доли третьей лобной извилины, нижней теменной и отчасти височной, поскольку их поражение даёт наиболее серьёзные нарушения высших психических процессов. Функциональная многозначность обусловлена при этом ещё и тем, что психическая функция связана не с механизмом или аппаратом как таковым, а с его динамически изменяющимся состоянием или, точнее, с протекающими в нём невродинамическими процессами в их сложной «исторической» обусловленности.
Полное разрешение всех противоречий между различными исследователями по вопросу о функциональной локализации может быть достигнуто лишь на основе генетической точки зрения. Степень дифференциации коры и распределения функций между различными её участками на разных ступенях развития различна. Так, у птиц, которых изучал М. Ж. П. Флуранс, отрицавший всякую локализацию, ещё никакой локализации в коре, по-видимому, не существует. У средних млекопитающих, у собак, кошек, как установили опыты И. П. Павлова и Л. Лучиани, которые признают некоторую локализацию, но очень относительную — с переслаиванием различных зон, некоторая локализация уже несомненно налицо; однако она ещё очень относительна: различные зоны переслаиваются. П. Флексиг, который утверждал, что в коре существуют особые проекционные зоны, т. е. зоны представительства различных рецепторных систем, связанных между собой расположенными между ними ассоциационными зонами, проводил свои исследования по преимуществу над человеческими эмбрионами. Различие результатов, к которым пришли эти исследователи по вопросу о локализации, объясняется, по-видимому, прежде всего различием тех объектов, которые они изучали.
Очевидно, неправильно механически переносить результаты, полученные в исследовании над одним животным, на других, стоящих на иных генетических ступенях, и обобщать эти результаты в общую теорию применительно к локализации функций в мозгу вообще. В этом (как правильно указывает Л. А. Орбели) заключается источник неправомерных споров и существенная причина расхождения разных точек зрения на локализационную проблему. Из того, что на низших ступенях эволюционного ряда нет локализации, нельзя делать вывода, что её нет и на высших: точно так же из её наличия на высших нельзя делать вывода о её наличии и на низших. Из того, что чувствительные зоны переслаиваются у собак, нельзя умозаключать, что это же имеет место на всех ступенях развития, в том числе и у человека, — так же как на основании того, что в коре человека выделяются относительно обособленные проекционные зоны, связанные расположенными между ними ассоциационными зонами, нельзя считать, что такова вообще структура мозга. Вопрос о функциональной локализации должен разрешаться по-разному для разных генетических ступеней — по-одному для птиц, по-другому для кошек и собак и опять-таки по-иному для человека.
В соответствии с той же основной генетической точкой зрения, нужно признать, что и для человека на данной ступени развития этот вопрос о локализации применительно к разным — генетически более древним и генетически более молодым — механизмам также решается по-разному: чем филогенетически древнее какой-либо «механизм», тем строже его локализация. Локализация в низших этажах нервной системы строже, чем в подкорке, в подкорке строже, чем в коре. В коре, в свою очередь, относительно примитивные «механизмы» в процессе филогенеза точнее закрепились за определёнными участками её; в осуществлении же высших генетически более поздних функций, сложившихся в процессе исторического развития человека, принимают участие очень многие или все «поля» коры, но различные поля, включаясь в работу целого, вносят в него различный вклад.
Физиология высшей нервной деятельности
Поскольку кора настолько существенна для психических процессов человека, особенно большое значение приобретает вопрос о том, каковы основные закономерности, определяющие её деятельность.
Первые основы для разрешения этого вопроса положили классические исследования И. П. Павлова. Они установили понятие условного рефлекса,как специфического типа деятельности коры.
В своём учении об условных рефлексах Павлов впервые создал подлинную физиологию коры больших полушарий головного мозга, т. е. того высшего отдела центральной нервной системы, функцией которого собственно является психика. В этом огромное значение павловского учения для научной психологии. Методика условных рефлексов является мощным и плодотворным методом объективного изучения психики.
Исходя из созданной им (намеченной уже Сеченовым) концепции об условно-рефлекторной деятельности, Павлов установил первые основные законы деятельности центральной нервной системы, или «высшей нервной деятельности». Таково правило нервного замыкания,которое устанавливает как общее явление в высшем отделе центральной нервной системы, что всякий сильно раздражаемый центр направляет к себе всякое другое более слабое раздражение, попадающее в то же время в эту систему; таким образом, пункт приложения этого раздражения и центр более или менее прочно связываются на определённое время при определённых условиях. Необходимым условием образования такой связи — условного рефлекса — является некоторое предшествование во времени слабого раздражения сильному.
Правило перехода клетки в тормозное состояние устанавливает, что если условный положительный раздражитель, вызывающий соответствующую условную реакцию, продолжает действовать некоторое время один, без дальнейшего сопровождения его безусловным раздражителем, то корковая клетка этого раздражителя переходит в тормозное состояние. Соответствующий раздражитель, систематически применяясь один, становится условным тормозным, отрицательным раздражителем: он обусловливает не процесс раздражения, а процесс торможения.
Правило или закон иррадиирования и концентрирования нервных процессов заключается в том, что процессы раздражения и торможения, возникнув в определённых пунктах коры под влиянием соответствующих раздражений, иррадиируют, распространяются по коре на большее или меньшее протяжение, а затем снова концентрируются, сосредоточиваются в ограниченном пространстве.
Наконец, правило или закон индукции нервных процессов устанавливает существование, наряду с явлениями раздражения и торможения, иррадиирования и концентрирования, переплетающегося с ними явления индукции противоположных процессов, т. е. усиление одного процесса другим, как последовательно на одном и том же пункте, так и одновременно на соседних пунктах.
Особое правило фиксирует зависимость величины эффекта от силы раздражения: правило предела силы раздражения устанавливает наличие для корковых клеток максимального раздражителя или предела безвредного функционального напряжения, за которым следует торможение. Торможение, вызываемое сверхмаксимальным раздражителем, искажает правило зависимости величины эффекта от силы раздражения,и сверхсильный раздражитель даёт эффект такой же или меньший, чем слабый; наступает так называемая уравнительная или парадоксальная фаза. [См. акад. И. П. Павлов, Двадцатилетний опыт объективного изучения высшей нервной деятельности (поведения) животных. «Условные рефлексы», М.—Л. 1938.]
Вся эта система правил или законов очерчивает определённый, ставшей благодаря работам Павлова классическим, тип деятельности коры. Этот новый тип условно-рефлекторной деятельности коры составляет физиологическую основу ассоциативных процессов.
Учение Павлова положило конец метафизической физиологии вульгарного ассоцианизма, который, проецируя ассоциацию в мозг, представлял её очень упрощённо, как «проторение путей». Раскрытые им закономерности возбуждения и торможения, иррадиации, концентрации нервных процессов и индукции вскрыли всю сложность реальных процессов, составляющих физиологический аспект таких в психологическом плане относительно элементарных явлений, как ассоциация.
Однако к одним только ассоциациям вся психическая деятельность человека в целом не сводима. Поэтому естественно возникает вопрос о специфической физиологической основе высших типов психической деятельности человека. Павлов сам указал на необходимость изучения новых законов того, что он назвал второй сигнальной системой, связанной с ролью речи в психике человека. Дать на этот вопрос ответ, хотя бы приближающийся к той классической чёткости, с какою Павлов разработал учение об условных рефлексах, остаётся нелёгкой задачей будущих исследований. Они должны завершить дело Павлова, разработав учение о физиологических механизмах высших форм сознательной деятельности человека.
Date: 2015-10-21; view: 556; Нарушение авторских прав