Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Структура ионного канала
|
|
Ион-селективный канал состоит из следующих частей (рис. 11): погруженной в бислой белковой части, имеющей субъединичное строение; селективного фильтра, образованного отрицательно заряженными атомами кислорода, которые жестко расположены на определенном расстоянии друг от друга и пропускают ионы только определенного диаметра; воротной части.
Ворота ионного канала управляются мембранным потенциалом и могут находиться как в закрытом состоянии (штриховая линия), так и в открытом состоянии (сплошная линия). Нормальное положение ворот натриевого канала - закрытое. Под действием электрического поля увеличивается вероятность открытого состояния, ворота открываются и поток гидратированных ионов получает возможность проходить сквозь селективный фильтр.
Если ион подходит по диаметру, то он сбрасывает гидратную оболочку и проскакивает на другую сторону ионного канала. Если же ион слишком велик по диаметру, как, например, тетраэтиламмоний, он не в состоянии пролезть сквозь фильтр и не может пересечь мембрану. Если же, напротив, ион слишком мал, то у него возникают сложности в селективном фильтре, на сей раз связанные с трудностью сброса гидратной оболочки иона.
Блокаторы ионных каналов либо не могут пройти сквозь него, застревая в фильтре, либо, если это большие молекулы, как ТТХ, они стерически соответствуют какому-либо входу в канал. Так как блокаторы несут положительный заряд, их заряженная часть втягивается в канал к селективному фильтру как обычный катион, а макромолекула закупоривает его.
Таким образом, изменения электрических свойств возбудимых биомембран осуществляется с помощью ионных каналов. Это белковые макромолекулы, пронизывающие липидный бислой, которые могут находиться в нескольких дискретных состояниях. Свойства каналов, селективных для ионов К+, Na+ и Са2+, могут по-разному зависеть от мембранного потенциала, что и определяет динамику потенциала действия в мембране, а также отличия таких потенциалов в мембранах разных клеток.
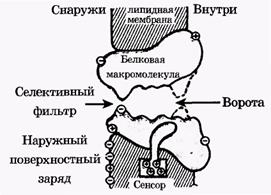
Рис. 11. Схема строения натриевого ионного канала мембраны в разрезе
В нервных волокнах сигналы передаются с помощью потенциалов действия, которые представляют собой быстрые изменения мембранного потенциала, быстро распространяющиеся вдоль мембраны нервного волокна. Каждый потенциал действия начинается со стремительного сдвига потенциала покоя от нормального отрицательного значения до положительной величины, затем он почти так же быстро возвращается к отрицательному потенциалу. При проведении нервного сигнала потенциал действия движется вдоль нервного волокна вплоть до его окончания. На рисунке показаны изменения, возникающие на мембране во время потенциала действия, с переносом положительных зарядов внутрь волокна вначале и возвращением положительных зарядов наружу в конце. В нижней части рисунка графически представлены последовательные изменения мембранного потенциала в течение нескольких 1/10000 сек, иллюстрирующие взрывное начало потенциала действия и почти столь же быстрое восстановление. Стадия покоя. Эта стадия представлена мембранным потенциалом покоя, который предшествует потенциалу действия. Мембрана во время этой стадии поляризована в связи с наличием отрицательного мембранного потенциала, равного -90 мВ. Фаза деполяризации. В это время мембрана внезапно становится высокопроницаемой для ионов натрия, позволяя огромному числу положительно заряженных ионов натрия диффундировать внутрь аксона. Нормальное поляризованное состояние в -90 мВ немедленно нейтрализуется поступающими внутрь положительно заряженными ионами натрия, в результате потенциал стремительно нарастает в положительном направлении. Этот процесс называют деполяризацией, В крупных нервных волокнах значительный избыток входящих внутрь положительных ионов натрия обычно приводит к тому, что мембранный потенциал «проскакивает» за пределы нулевого уровня, становясь слегка положительным. В некоторых более мелких волокнах, как и в большинстве нейронов центральной нервной системы, потенциал достигает нулевого уровня, не «перескакивая» его. Фаза реполяризации. В течение нескольких долей миллисекунды после резкого повышения проницаемости мембраны для ионов натрия, натриевые каналы начинают закрываться, а калиевые — открываться. В результате быстрая диффузия ионов калия наружу восстанавливает нормальный отрицательный мембранный потенциал покоя. Этот процесс называют реполя-ризацией мембраны. Для более полного понимания факторов, являющихся причиной деполяризации и реполяризации, необходимо изучить особенности двух других типов транспортных каналов в мембране нервного волокна: электроуправляемых натриевых и калиевых каналов. Электроупавляемые натриевые и калиевые каналы. Необходимым участником процессов деполяризации и реполяризации во время развития потенциала действия в мембране нервного волокна является электроуправляемый натриевый канал. Электроуправляемый калиевый канал также играет важную роль в увеличении скорости реполяризации мембраны. Оба типа электроуправляемых каналов существуют дополнительно к Na+/K+ -насосу и каналам К*/Na+-утечки. Электроуправляемый натриевый канал. В верхней части рисунка показан электроуправляемый натриевый канал в трех различных состояниях. Этот канал имеет двое ворот: одни вблизи наружной части канала, которые называют активационными воротами, другие — у внутренней части канала, которые называют инактивационными воротами. В верхней левой части рисунка изображено состояние этих ворот в покое, когда мембранный потенциал покоя равен -90 мВ. В этих условиях активационные ворота закрыты и препятствуют поступлению ионов натрия внутрь волокна. Активация натриевого канала. Когда мембранный потенциал покоя смещается в направлении менее отрицательных значений, поднимаясь от -90 мВ в сторону нуля, на определенном уровне (обычно между -70 и -50 мВ) происходит внезапное конформационное изменение актива-ционных ворот, в результате они переходят в полностью открытое состояние. Это состояние называют активированным состоянием канала, при котором ионы натрия могут свободно входить через него внутрь волокна; при этом натриевая проницаемость мембраны возрастает в диапазоне от 500 до 5000 раз. Инактивация натриевого канала. В верхней правой части рисунке показано третье состояние натриевого канала. Увеличение потенциала, открывающее активационные ворота, закрывает инактивационные ворота. Однако инактивационные ворота закрываются в течение нескольких десятых долей миллисекунды после открытия активационных ворот. Это значит, что конформационное изменение, приводящее к закрытию инактивационных ворот, — процесс более медленный, чем конформационное изменение, открывающее активационные ворота. В результате через несколько десятых долей миллисекунды после открытия натриевого канала инактивационные ворота закрываются, и ионы натрия не могут более проникать внутрь волокна. С этого момента мембранный потенциал начинает возвращаться к уровню покоя, т.е. начинается процесс реполяризации. Существует другая важная характеристикая процесса инактивации натриевого канала: инактивационные ворота не открываются повторно до тех пор, пока мембранный потенциал не вернется к значению, равному или близкому к уровню исходного потенциала покоя. В связи с этим повторное открытие натриевых каналов обычно невозможно без предварительной реполяризации нервного волокна.
Потенцил Действия
1. Объясняется поворотом диполя на 1800.
2. Теория альтерации Германа. При возбуждении возникает избыток кислых продуктов, которые несут отрицательны заряд, что приводит к разности потенциалов между возбужденным и невозбужденым учаском.
3. Мембранная теория Берштейна. В возбужденном участке мембраны резко увеличивается проницаемость для всех ионов, концентрации ионов смешиваются и участок становится электронейтральным.
4. Ходжкин и Хаксли. Рост проницаемости мембраны для ионов в месте воздействия. При возбуждении электропроницаемость мембраны увеличивается примерно в 500 раз. Max увеличивается проницаемость мембраны для Na+ (отсюда Na-теория ПД). Na+ свободно проходит внуть клетки. При возбужедении электро-химическое равновесие определяется потенциалом Na+. Равновесный потенциал для К+ = –97 мВ, для Na+ = +50 мВ. При возбужедении мембрана перезаряжается. Положение обратной активации и инактивации Na+-каналов, Na+-канал может активироваться (открываться) при определенных значениях потенциала. Причина активации Na+-каналов – деполяризация мембраны, чем больше деполяризация, тем больше проницаемость мембраны для Na+. Зависимость близка к линейной в подкор уровне; как только мембрана достигнет критического уровня деполяризации – зависимость нелинейная, лавинообразный вход Na+ в клетку.
1). Для объяснения реполяризации используется положение об инактивации Na+-каналов. При приближении потенциала мембраны к равновесному для Na+, Na+-каналы инактивируются и посупление Na в клетку прекращается. К графику: в основе регенеративный процесс (сам себя поддерживающий), развивающийся по принципу обратной связи.
2). Рост К+ проницаемости мембраны. Не столь значителен, как для Na+ (в 5-15 и 500 раз соответственно). Проницаемость для К+ развивается медленнее, чем для Na+. Ионы К+ в этой ситуации будут выходить наружу и выносить заряд.
3). Механизм активного транспорта, представленный K-Na-насосом. 3 Na+ внутрь и 2 К+ наружу.
Эксперименты Ходжкина и Хаксли.
Гигантский аксон кальмара. Из внеклеточной среды были удалены 2/3 Na+. При этом амплитуда ПД снизилась» на 50%. Замена внутриклеточного Na+ на другие ионы приводит к некоторому росту ПД. Замена Ѕ внутриклеточного К+ на Na+приводит к значительному снижению ПД.
Метод фиксации потенциала
метод Петч-Клемпинга. С его помощью можно зафиксировать на длительное время значение мембранного потенциала на любом желаемом уровне. Это делается с помощью внешнего генератора напряжения
Суммарные мембранные токи при ПД
1. Подпороговая область:
Слабое изменение мембранного потенциала, суммарный ионный ток направлен от клетки наружу, так как поток К+, выходящий из клетки, уже усиливается из-за удаления мембранного потенциала от равновесного потенциала для К+. Входящий ток Na+ еще слаб, так как рост Na+-проницаемости пока невелик. Однако с развитием деполяризации Na-ый поток постепенно нарастает.
2. Критический уровень деполяризации:
В этот момент суммарный ионный ток через мембрану равен нулю, так как встречные токи ионов Na+ и К+ уравновешивают друг друга. Даже небольшая дальнейшая деполяризация приводит к росту входа Na+-тока в сотни раз.
3. Во время фазы деполяризации резко увеличивается Na+-проницаемость и суммарный мембранный ток, направленный внутрь клетки. Выходящий К+-ток растет медленнее и становится заметным только к моменту пика потенциала.
4. Фаза реполяризации:
В момент пика потенциала большинство Na+-каналов инактивированны, а К+-ток max. Поэтому суммарный мембранный ток – выходящий.
Кальциевая теория активации и инактивации Na+-каналов
В состоянии покоя у наружного отверстия Na+-канала находится Са2+, который электростатически тормозит проникновение Na+ в канал. При возбуждении наружная поверхность мембраны заряжена отрицательно, при этом Са2+ уходят со своих мест, вход открывается и Na+ входит в клетки.
Инактивация: по ходу деполяризации узкие Na+-каналы могут закупориваться Na+. Во многих каналах есть воротные белки (могут менять свое местоположение под влиянием изменения потенциала). В состоянии покоя активационный белок закрыт, а инактивационный открыт. При возбуждении открывается активационный белок в момент закрывания инактивационного белка. В конце реполяризации белки так же закрываются и потом открываются (исходное состояние).
Передача возбуждения по нервным волокнам
В начале 30х годов ХХ в. Хилл. 1932 г. "Химическая волна проведения в нервах". Хилл использовал разные нервы, но преимущественно краба. Даже в состоянии покоя в единицу времени вырабатывается некоторое количество тепла. Это тепло было названо теплопродукция покоя. Когда в нервном волокне возбуждение – теплопродукция возбуждения (ТВ), она делится на 2 фазы:
1. Начальная ТВ, которая составляет 2-3% от всей ТВ и приходится непосредственно на период возбуждения.
2. Задержанная ТВ» 97% всей ТВ. Если подать серийный импульс на нерв краба, то задержанную ТВ можно зафиксировать в течение 25-30 минут. Возбуждения в тканях уже нет, но ТВ имеет место.
3. Утечка тепла при работе Na.
Хилл разрабтал чувствительную теплоэлектрическую методику, которая позволяла фиксировать теплообразование в течение 20 мс. Эксперименты при О0 С. Начальную фазу теплопродукции делили на 2 периода: позитивная и негативная начальная теплопродукция. При О0 С для нерва краба позитив в начальные 20 мс = 14 мк кал. В течение последующих 150 мс» 85% тепла поглощается нервной тканью обратно (12 мк кал).
Позитивная начальная теплопродукция: причина: химические процессы, обуславливающие изменение проницаемости мембраны. При возбуждении в клетку поступает Na+ и смешивается с К+ и наоборот. Должно образовываться тепло. Это тепло покрывает до 50% позитивной начальной теплопродукции.
Негативная начальная теплопродукция: химические реакции в этот период могут быть эндотермическими. Негативная теплопродукция не является обязательной.
Проведение возбуждения
В 1885 г. Герман предложил теорию малых токов. Осуществляется последовательно между участками волокна. В участке, соседнем с возбужденным будет наблюдаться выход электрического тока.
Кабельная теория нервного волокна: нервное волокно внутри содержит проводящую среду, оболочка невного волокна имеет слой, который плохо проводит возбуждение. Нервное волокно омывается внеклеточной жидкостью, которая проводит электрический ток.
Эквивалентная электрическая схема нервного волокна
В состоянии покоя внутриклеточная среда имеет избыточный отрицательный заряд. Сила тока меняется с расстоянием от возбужденного сегмента, декремента.
Факторы, определяющие скорость распространения возбуждения по нервному волокну
1. Пространственная константа определяет величину декремента деполяризации, l - пространственная константа.
2. Коэффициент надежности, соотношение между амплитудой ПД, критической энергией и ПП. S=ПД/(Екр–ПП), ПД=120мВ, ПП=–70мВ, Екр=–55мВ Þ S=8. Чем больше S, тем быстрее проведение.
3. Временная константа t мембр. При возбуждении мембраны меняется заряд. Длительность перезарядки мембраны.tмембр=Rm*Cm. Чем больше t мембр, тем ниже С. Vраспр=S*l/tмембр.
Механизм распространения возбуждения
Возбуждение охватывает последовательно все отделы нервного волокна. R наруж влияет на скорость распространения возбуждения. В экспериментрах Ходжкина изменили внеклеточную среду на масло, которое имеет большее сопротивление, объем снизился на 30-50%. Эксперимент: нерв помещается на параллельные пластинки из серебра, замыкают с помощью ртутной ванночки, объем проведеним растет на 16-30%. Была подтверждена теория местных токов для безмякотных волокон. В мякотных нерных волокнах механизм проведения другой. Миелин имеет рост сопротивления и снижение емкости, миелиновая оболочка прерывается перехватами Ранвье – сальтаторное проведение. R на 1 см2 поверхности в перехвате Ранвье = 10-20 Ом, в миелиновой оболочке = 0,003-0,005 Ом. Петли тока в миелиновых нервных волокнах выходят через невозбужденый перехват Ранвье, находящийся спереди от возбужденного. Эксперименты Тасаки.
1. Электроды стоят на миелине, два выходящих тока (это токи, выходящие из последующего и предыдущего перехвата Ранвье. Входящий ток не регистрируется.
2. Средний электрод на перехвате. Появляется входящий ток.
- Электрическая активность органов
функционирование живых клеток сопровождается возникновением трансмембранных потенциалов - биопотенциалов. Клетки, образуя целостный орган, формируют сложную картину его электрической активности. Она определяется как электрической активностью отдельных клеток, так и взаимодействием между ними, устройством самого органа, неоднородностью структуры этого органа, процессами регуляции в нем и целым рядом других причин.
Электрическая активность в большой степени отражает функциональное состояние клеток, тканей и органов. Регистрация и анализ электрической активности позволяют проводить биофизические и медико-биологические исследования с целью изучения работы органов и проведения клинической диагностики.
лектрическая активность органов и тканей обладает важной информацией о происходящих в них физиологических и патологических процессах. Для получения этой информации следует зарегистрировать электрическую активность.Необходимо знать электрические характеристики «генераторов» (органов и тканей), так как без учета этих характеристик невозможно неискаженно зарегистрировать биоэлектрические процессы. Всякий электрический генератор, в том числе электрически активный орган или ткань, характеризуется следующими параметрами: а) характером изменения ЭДС во времени (диапазоном изменения амплитуд ЭДС, частотным диапазоном и формой); б) внутренним сопротивлением. Прежде чем рассмотреть эти параметры применительно к органам и тканям, необходимо остановиться на некоторых терминах. При изучении незатухающих синусоидальных колебаний под их амплитудой понимают наибольшее отклонение колеблющейся величины от среднего значения. Кривые биоэлектрической активности органов и тканей значительно отличаются от синусоиды и поэтому под амплитудой в электрографии условно понимают либо размах колебаний между пиками, либо величину отклонения кривой от средней линии, что, как правило, оговаривают. Под периодом незатухающих синусоидальных колебаний - Т понимают время, в течение которого совершается одно полное колебание; частотой - fназывается число периодов за одну секунду
f = 1/Т (1) Биоэлектрические процессы можно условно разделить на две группы - квазипериодические (электрическая активность сердца, a=ритм электрической активности мозга) и апериодические (электрическая активность мышц и др.). Но даже в квазипериодических биоэлектрических процессах период не остается неизменным и поэтому под частотой колебаний нужно понимать их среднюю частоту за какое-то определенное время. Какими же характеристиками обладают ткани и органы человека и животных, если их рассматривать как электрические генераторы?При возбуждении клетки - элементарного электрического генератора - ее возбужденная часть становится электроотрицательной относительно невозбужденной части и разность потенциалов достигает 50 мв. Клетки в органах и тканях человека и животных соединены параллельно, поэтому суммарная ЭДС возбужденного органа должна была быть того же порядка, что и ЭДС клетки. Однако суммарная электрическая активность клеток, регистрируемая с органа, всегда бывает ниже электрической активности отдельной клетки. Это объясняется тем, что в органе элементарные электрические генераторы - клетки, имеющие относительно большое внутреннее сопротивление, шунтируются сравнительно малым сопротивлением межклеточной жидкости, что приводит к ощутимому снижению разности потенциалов, развиваемой возбужденным органом. При регистрации биоэлектрической активности мышц и сердца человека с помощью электродов, наложенных на кожу, амплитуда колебаний не превышает единиц милливольт, а амплитуда электрической активности мозга - сотом микровольт. Таким образом, можно сделать вывод, что амплитуды колебаний бионапряжений весьма малы и составляют тысячные и стотысячные доли вольта. Некоторые виды биоэлектрической активности тканей и органов являются апериодичными процессами сложной формы (например, электрическая активность мышц). Для того чтобы зарегистрировать эти процессы неискаженно, необходимо, чтобы электрографическая установка обеспечивала запись совершенно определенной полосы частот. Именно в этом смысле говорят, что, например, суммарная биоэлектрическая активность мышцы занимает диапазон частот от 1 до 1000 Гц. Если учесть, что некоторые биоэлектрические процессы изменяются весьма медленно (кожные потенциалы), а для неискаженного воспроизведения других (электрическая активность одиночного мышечного волокна) требуется регистрация колебаний в десятки тысяч герц, то можно считать, что биоэлектрические процессы человека занимают диапазон от постоянных напряжений и инфранизких до низких частот включительно. При регистрации биоэлектрических процессов человека, внутренним сопротивлением эквивалентного электрического генератора, например, мышцы, является междуэлектродное сопротивление, включающее в себя сопротивление кожи, ряда других тканей и сопротивление органа, электрическая активность которого регистрируется. Оно зависит от ряда факторов (сила и форма тока, площадь электродов, качество обработки кожи, температура воздуха и др.) и достигает большой величины. На требования к электрографическим установкам, естественно, влияют характеристики органов и тканей как электрических генераторов. Так, низкая амплитуда биоэлектрических процессов органов и тканей человека ведет к тому, что электрографические установки должны обладать весьма высокой чувствительностью, а их усилители - высоким коэффициентом усиления. Эти установки должны также обеспечивать неискаженную регистрацию постоянных и медленно меняющихся разностей потенциалов, инфранизких и низкочастотных колебаний потенциалов. И, наконец, для того чтобы электрограмма, записанная с помощью электродов, наложенных на кожу человека, была тождественна электрической активности исследуемого органа, входное сопротивление установки должно быть во много раз больше, чем междуэлектродное сопротивление. Электрически активный орган окружен тканями, являющимися объемным проводником. Разность потенциалов, продуцируемая органом, вызывает в окружающих его тканях биотоки, и, следовательно, в последних создаются разности потенциалов, повторяющие все изменения ЭДС электрически активного органа. Для того чтобы составить себе представление, какие разности потенциалов и потенциалы продуцируют электрически активный орган в окружающих его тканях, необходимо рассмотреть упрощенную модель.
- Физические основы электрокардиографии
Каждая клетка сердечной мышцы создаёт электрическое поле, которое имеет характеристики, подобные в общих чертах характеристикам электрического поля других типов мышечных клеток. Но потенциал действия (ПД) сердечных клеток отличается от ПД клеток поперечнополосатых мышц своей формой и длительностью. Электрическое поле сердца в целом образуется наложением электрических полей отдельных клеток. Изменения электрического поля сердца происходят при деполяризации и реполяризации мембраны клеток сердца (Рис.4). Эти изменения достаточны, чтобы создать изменения разности потенциалов между различными точками поверхности тела и чтобы обнаружить указанные изменения на большом расстоянии от их источника.
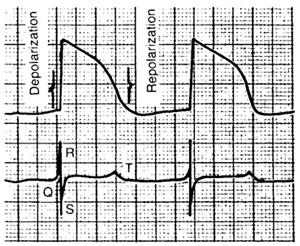 Рис. 4. Потенциал действия сердечной клетки и соответствующие изменения электрического поля сердца
Рис. 4. Потенциал действия сердечной клетки и соответствующие изменения электрического поля сердца
Графическая запись электрического потенциала, созданного возбуждением клеток сердца, называется электрокардиограммой (ЭКГ). Таким образом, ЭКГ характеризует возбуждение сердца, но не его сокращения.
Впервые электрокардиограмма была записана голландским физиологом Эйнтховеном посредством сравнительно простого инструмента струнного гальванометра. В настоящее время для записи ЭКГ используют специальные электронные приборы, называемые электрокардиографами. Амплитуда электрического потенциала записанного с поверхности тела может быть менее 1 мВ. Следовательно, перед записью потенциал должен быть усилен с помощью устройства, называемого усилителем. Электрокардиограф включает также высокочастотное сито, не пропускающее медленные изменения электрического потенциала, и калибратор, который генерирует электрические импульсы 1 мВ, что необходимо для расчета амплитуды зубцов электрокардиограммы.
Date: 2015-09-24; view: 1286; Нарушение авторских прав