Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Физиологические особенности гладких мышц. Гладкомышечные клетки делят на два типа: сосудистого и висцерального
|
|
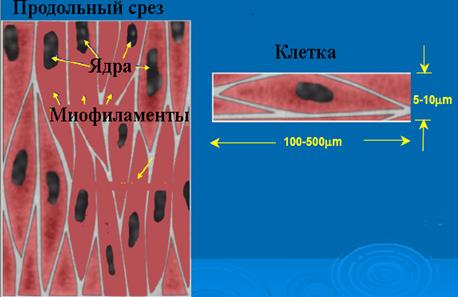
Гладкомышечные клетки делят на два типа: сосудистого и висцерального. Гладкомышечная клетка сосудистого типа имеет размеры 20´5-10 мкм, у клеток висцерального типа эти параметры равны 100-200´10-20 мкм. Плазматическая мембрана гладкомышечной клетки имеет особые впячивания - кавеолы, благодаря которым площадь мембраны увеличивается на 70%. Между рядами кавеол располагаются электронно-плотные тяжи, связанные с актиновыми протофибриллами. Снаружи плазматическая мембрана покрыта базальной мембраной и отделена от нее электронно-светлой областью. Комплекс базальной и плазматической мембраны называют сарколеммой. Ядро располагается в средней утолщенной части клетки и не имеет особенностей по сравнению с клетками других типов. СПР состоит из гранулярной и гладкой части, гранулярная часть лежит у ядра, гладкая - вблизи плазмалеммы и часто тесно контактирует с кавеолами. Гладкомышечные клетки лежат в виде параллельно ориентированных пучков, расстояние между ними заполнено коллагеновыми и эластическими волокнами, фибробластами, питающими магистралями. Между клетками имеются тесные контакты: 1) нексусы - щели ~2-3 нм, имеющие ионные каналы и 2) десмосомоподобные контакты - щели 20-60 нм, заполненные электронно-плотным веществом, переходящим в базальную мембрану, выполняют механическую функцию, связывая клетки.
Основу сократительного аппарата составляют миозиновые и актиновые протофибриллы. В ГМК актиновых протофибрилл намного больше, чем в поперечно-полосатом мышечном волокне. Соотношение актин/миозин = 5:1.
Миозиновые протофибриллы имеют толщину 14 нм, длину 2,6 мкм - т.е. заметно больше, чем в скелетных мышцах, и состоят из миозиновых молекул. В миоците миозин агрегирует в толстые протофибриллы, с образованием той же пространственной структуры, как и миозин скелетных мышц.
Актиновые протофибриллы имеют удлиненную форму, гексагональны на срезе и представляют собой агрегаты актина, тропомиозина и кальдесмона. Глобулы G-актина полимеризуются в 2 цепи F-актина, образующие видоизмененный остов молекулы фибриллы. Тропомиозина в ГМК в 7 раз меньше, чем в скелетных мышцах, они расположены в желобках каркаса актиновой глобулы, контактируя с 7 глобулами F-актина. Тропонин скелетных мышц в ГМК отсутствует.
Помимо описанных 2 типов протофибрилл в ГМК есть промежуточные фибриллы, весьма стойкие к различным денатурирующим агентам. Эти протофибриллы образуют трехмерную сеть, крепящуюся к плотным тельцам десмосом и межкавеолярных тяжей, которая служит сократительным остовом (своеобразной внутриклеточной «точкой опоры», к которой и прилагается сила сокращения).
В ГМК протофибриллы, в отличие от скелетной мышцы, не пакуются в миофибриллы, саркомера как такового нет, роль Z-пластинок выполняют плотные тельца. В ГМК отсутствует Ca++-тропониновая система активации сокращения, вместо нее ведущая роль принадлежит системе фосфорилирования. Возрастание концентрации внутриклеточного кальция [Cai++] и взаимодействие его с кальмодулином приводит к повышению аффинности (сродства) комплекса Са++-кальмодулин к киназе L-цепи миозина (ее каталитической единицы). Происходящее при этом фосфорилирование легкой цепи миозина вызывает активацию актинзависимой Mg++-АТФ-азы миозина и генерацию сокращения. Ингибиторами фосфорилирования L-цепи миозина служат цАМФ-зависимые протеинкиназы, которые в отсутствие Са++ фосфорилируют 2 центра киназы L-цепей и снижают тем самым их аффинность к комплексу Са++-кальмодулин, а это, в конечном итоге, уменьшает фосфорилирование L-цепей. Помимо фосфорилирующей системы сокращение ГМК регулирует независимый Са++-чувствительный механизм. При этом фосфорилирующая система контролирует скорость циклирования поперечных мостиков (присоединения-отсоединения миозина к актину) в фазу развития напряжения, а Са++-чувствительный механизм - в фазу тонуса.
Электрофизиологические явления в ГМК. Если в анатомическом смысле гладкомышечная ткань представляет собой слаженный ансамбль отдельных клеток, то в функциональном плане - это электрический синцитий. ГМК имеет ряд электрофизиологических особенностей: 1) клетка не способна (за небольшим исключением) генерировать «полноценный» потенциал действия (ПД), вместо этого на мембране возникает локальная деполяризация; 2) электротонические потенциалы способны распространяться на большие расстояния, не затухая. Распространение электротонического потенциала идет по нексусам, обладающих чрезвычайно низким сопротивлением.
Потенциал покоя (ПП) ГМК. Проводимость мембраны ГМК для одновалентных ионов составляет pK+:pNa+:pCl- = 1:0,2:0,6-0,7. Однако, в генерации ПП клетки играют ведущую роль не только эти ионные токи, но и электрогенный компонент Na+-K+ насоса (что в меньшей степени характерно для ПП скелетного мышечного волокна). ПП клетки колеблется в пределах -50¸-70 мВ, тогда как пороговый потенциал (при котором включаются потенциалзависимые Са++ каналы) составляет -30¸-35 мВ. Основной вклад в ПП ГМК вносят ионы К+ (Ko+= 2-3 мМ, Кi+= 160 мМ). Ионы Na+ и Cl- оказывают на величину ПП клетки лишь мягкое «регулирующее» действие.
Потенциал действия ГМК. Спонтанная активность ГМК проявляется в виде медленной волны деполяризации (15-20 мВ, длительность 250-400 мс) и предшествующего ей пикового потенциала (25-50 мВ, длительность 50 мс). Часто вслед за первым идет второй пиковый потенциал, чем больше пауза после первого пикового потенциала, тем больше амплитуда и меньше длительность второго пика. Второй пиковый потенциал в отличие от первого никогда не прерывает развитие медленной волны деполяризации. Спонтанная активность ГМК имеет миогенную природу, за счет пейсмекерной активности ГМК, но в целом зависит от уровня ПП (чем он ниже - тем меньше активность). Ионная природа спонтанной активности обусловлена изменением концентрации ионов Са++ у наружной стороны мембраны. Удаление Са++ вначале усиливает спонтанную активность, а затем угнетает пиковый, и далее, медленный ее компонент. Увеличение Сао++ сопровождается снижением частоты спонтанных разрядов и увеличением их амплитуды. Ионы Са++ обеспечивают фазу деполяризации ПД и медленной волны, тогда как ионы К+ и выходящий К+ ток - фазу реполяризации. ПД ГМК артериол зависит от мощности их симпатической иннервации, чем она выше, тем больше ПД генерируется.
Роль Са++ в сопряжении возбуждения и сокращения. В отличие от скелетных мышц, где функцию сопрягающего агента между мембраной и актомиозиновым комплексом выполняет Т-система и СПР, в ГМК эти образования либо отсутствуют (Т-система), либо их недостаточно (СПР составляет примерно 2-7% объема цитоплазмы). В ГМК функцию сопряжения выполняют, очевидно, кавеолы плазматической мембраны ГМК и приближенные к ним участки СПР.
Важную роль в сопряжении процесса возбуждения и сокращения играют ионы Са++, которые снимают тормозящее воздействие тропонина на актин-миозиновое взаимодействие и запускают сокращения. В ГМК исходная концентрация Са++ составляет 2,0-2,8´10-7 моль/л, повышаясь до 3,8´10-6 моль/л на максимуме сокращения. Источниками Са++ для ГМК служат: 1) ионизированный Са++ внутриклеточной среды; 2) Са++ токи плазматической мембраны через 2 класса Са++-каналов и Na+-неселективные каналы; 3) внутриклеточные депо Са++ - митохондрии и СПР, внутренняя поверхность плазматической мембраны с ее Са++-связывающими белками. В отличие от скелетной мышцы, в ГМК ведущая роль принадлежит не СПР, а плазмалеммальным токам, т.к. благодаря относительно малому объему каждой клетки по сравнению с поперечно-полосатым мышечным волокном диффузионное расстояние плазмалемма - миофибрилла падает.
Date: 2015-09-19; view: 732; Нарушение авторских прав