Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Приложение 709261417. Энергетика мышечного сокращения
|
|
Единственным прямым (непосредственным) источником энергии для мышечного сокращения служит аденозинтрифосфат (АТФ). При гидролизе АТФ образуется аденозиндифосфат (АДФ), отщепляется фосфатная группа и выделяется около 10 ккал свободной энергии[Мф39].
Ресинтез АТФ
Для того чтобы мышечные волокна могли поддерживать сколько-нибудь длительное сокращение, необходимо постоянное восстановление АТФ с такой же скоростью, с какой он расщепляется (расходуется).
Энергия для восстановления АТФ образуется за счет питательных («горючих») веществ — углеводов, жиров, белков. При распаде (полном или частичном) этих веществ часть энергии, запасенная в их химических связях, освобождается. Эта свободная энергия обеспечивает связывание АДФ и фосфата с образованием АТФ.
Ресинтез АТФ в мышце может происходить двумя основными путями — анаэробным (без участия кислорода) и аэробным (при участии кислорода). Если энергетическое обеспечение работы осуществляется в основном анаэробным путем, такая работа называется анаэробной, а если преимущественно аэробным путем — аэробной работой.
Для образования и использования АТФ в качестве непосредственного источника энергии в сокращающейся мышце могут действовать три химические (энергетические) системы:
1) фосфагенная (АТФ-КФ-система)
2) гликолитическая (лактацидная)
3) окислительная (кислородная)
Фосфагенная и гликолитическая работают по анаэробному пути, окислительная — по аэробному.
Системы отличаются друг от друга по энергетической ёмкости и мощности[Мф40].
Энергетическая ёмкость системы — это максимальное количество образуемой энергии, или максимальное количество АТФ, которое может_ресинтезироваться за счет_энергии рассматриваемой системы (табл. 210210903).
Энергетическая мощность системы — это максимальное количество энергии, выделяющейся в единицу времени, или максимальное количество АТФ, образующейся в единицу времени за счет энергии рассматриваемой системы (табл. 210210903).
Емкость энергетической системы ограничивает максимальный объем работы, который может быть выполнен за счет данной системы.
Мощность системы лимитирует предельную интенсивность работы, выполняемой за счет энергии данной системы.
Таблица 210210903
Максимальная мощность и ёмкость трех энергетических систем (примерные величины из расчета на 20 кг активной мышечной массы*)
| Системы | Максимальная мощность, моль АТФ/мин | Емкость, общее число молей АТФ | |
| Фосфагенная | 3,6 | 0,5 | |
| Гликолитическая | 1,2 | 1,2 | |
| Окислительная | Окисление гликогена и глюкозы | 0,8 | |
| Окисление жиров | 0,4 |
* У «стандартного» мужчины весом 70 кг мышечная масса составляет около 40 % веса тела, т.е. примерно 30 кг. При наиболее типичных видах мышечной деятельности (локомоциях) участвует не более 2/3 всей мышечной массы. Общая масса сокращающихся мышц — активная мышечная масса — не превышает обычно 20 кг.
Степень участия трех систем в энергообеспечении (ресинтезе АТФ) зависит от силы и продолжительности мышечных сокращений, т.е. от мощности и длительности работы, а также от условий работы мышц, особенно от уровня их обеспечения кислородом.

Фосфагенная энергетическая система
К этой группе относятся высокоэнергетические фосфатные соединение АТФ и креатинфосфат (КрФ).
Восстановление (ресинтез, регенерация) АТФ осуществляется во время мышечных сокращений почти мгновенно, как только он расщепляется до АДФ.
КрФ — первый энергетический резерв мышцы, действующий как немедленный источник восстановления АТФ.
Конечными продуктами расщепления КрФ являются креатин и свободный фосфат. Энергия от расщепления КрФ немедленно и непосредственно используется для ресинтеза АТФ.
При отщеплении фосфатной группы от КрФ освобождается в среднем 10,5 ккал[V.G.41] [Мф42].
Мощность фосфагенной системы наибольшая по сравнению с другими системами (таблица 210210903). Максимальная скорость энергообразования, выраженная в количестве АТФ, ресинтезируемого в единицу времени за счет активности фосфагенной системы, примерно в 3 раза превышает максимальную мощность гликолитической системы и в 4—10 раз — максимальную мощность кислородной системы. Поэтому фосфагенная система играет решающую роль в энергообеспечении работ предельной мощности (спринтерский бег, кратковременные мышечные усилия «взрывного характера» — при подъеме штанги, метаниях, прыжках и т. п.), осуществляемых с максимально возможными по силе и скорости (мощности) сокращениями мышц.
Емкость фосфагенной системы невелика, так как запасы АТФ и КФ в мышце весьма ограниченны (таблица 210210903). Емкость фосфагенной системы, оцениваемая запасом АТФ в 20 кг мышц и количеством АТФ, которое может ресинтезироваться за счет КрФ, содержащегося в этой же мышечной массе, составляет всего около 0,5 М АТФ, т.е. около 5 ккал.
Поэтому работа с предельной мощностью, которая обеспечивается почти исключительно анаэробной фосфагенной системой, может продолжаться с энергетической точки зрения не более нескольких секунд. Например, при спринтерском беге энергетический запрос работающих мышц может достигать около 1 ккал/с. Поскольку максимальное количество энергии, которое может обеспечить фосфагенная система, составляет около 5 ккал, такая работа теоретически может продолжаться за счет фосфагенной системы не более 5 с.
Если выполняется работа большей продолжительности, её энергетическое обеспечение не может происходить только за счет наличных запасов мышечных фосфагенов (АТФ и КрФ).
Гликолитическая энергетическая система
В основе деятельности этой системы, обеспечивающей ресинтез АТФ и КФ, лежит цепь химических реакций анаэробного расщепления гликогена (и глюкозы). Совокупность этих реакций называется анаэробный гликогенолиз, или гликолиз (или цикл Эмбден — Мейерхофа). В результате таких реакций образуется молочная кислота — лактацид. Отсюда другое название этой энергетической системы — лактацидная. В результате гликолиза освобождается энергия, которая позволяет ресинтезировать АТФ.
Энергетическими субстратами лактацидной мышечной системы служат углеводы, главным образом гликоген, запасенный в мышечных клетках, и в меньшей степени — глюкоза. Молекула гликогена — это цепочка молекул глюкозы. При расщеплении молекулы гликогена от нее последовательно отщепляются молекулы глюкозы (глюкозные единицы). В мышечные клетки поступает также глюкоза из крови. В свою очередь, глюкоза попадает в кровь из печени. Анаэробное расщепление гликогена (гликогенолиз) в мышечных клетках имеет энергетическое преимущество перед гликолизом (анаэробным расщеплением глюкозы), так как каждая глюкозная единица из молекулы мышечного гликогена в процессе гликолиза дает энергию для образования 3 молекул АТФ, а каждая молекула глюкозы — только для 2 молекул АТФ.
В результате анаэробного гликогенолиза из каждой глюкозной единицы (глюк. ед.) образуются 2 молекулы молочной кислоты. Химические реакции анаэробного гликогенолиза (гликолиза) происходят исключительно в саркоплазме, где находятся все ферменты, катализирующие гликолиз (гликолитические ферменты).
Анаэробный гликогенолиз развертывается практически с начала мышечной работы, но достигает максимальной мощности (наибольшей скорости образования молочной кислоты) лишь через 30—40 с. Поэтому наибольшие концентрации молочной кислоты в мышцах (и в крови) обнаруживаются лишь при работах, которые продолжаются более 20—30 с. При более кратковременной работе энергетическая роль лактацидной системы относительно меньше.
Мощность лактацидной системы мышц велика. При работах очень большой мощности, которые могут продолжаться лишь несколько десятков секунд, скорость расходования гликогена в мышцах достигает 10 мМ глюк.ед/кг веса мышцы в минуту или около 0,2 г/мин (рис[V.G.43].).
Определение содержания и расходования в мышцах гликогена и других энергетических субсгратов у человека стало возможным благодаря использованию метода игольчатой биопсии — взятию малой порции мышечной ткани с помощью специальной иглы (Д.Бергстрём) до работы и в разные периоды ее выполнения, а также после работы.
Таким образом, скорость расщепления гликогена в активной мышечной массе в 20 кг равна 200 мМ глюк. ед/мин. Если расщепление гликогена идет анаэробным путем, то при этом образуется 400 мМ молочной кислоты в минуту. Образование каждой молекулы молочной кислоты из гликогена обеспечивает ресинтез 3 молекул АТФ. Следовательно, при образовании 400 мМ молочной кислоты ресинтезируется 1,2 М АТФ/мин. Такова примерная максимальная мощность лактацидной системы (см. табл. 2).
Мощность лактацидной системы в 1,5 раза выше, чем кислородной, но примерно в 3 раза ниже мощности фосфагенной системы (табл. 210210903).
Лактацидная система играет решающую роль в энергетическом обеспечении работ очень большой мощности, которые могут продолжаться от 20 с до 1—2 мин и связаны с сильными мышечными сокращениями, требующими большой скорости расщепления и соответственно ресинтеза АТФ. К таким видам мышечной работы относятся, например, бег на дистанциях от 200 до 800 м, плавание на дистанциях от 50 до 200 м и т.д.
По мере дальнейшего уменьшения мощности выполняемой работы (увеличения её продолжительности) энергетическая роль лактацидной системы снижается.
Лактацидная система функционирует в тех случаях, когда сокращающиеся мышцы испытывают недостаток в снабжении кислородом. Такие условия возникают:
при работах очень большой мощности,
в самом начале любой работы когда снабжение мышц кислородом отстает от потребности в нем [p],
при статическом сокращении мышц даже небольшой силы (превышающей 20% от максимальной).
При статическом сокращении мышц даже небольшой силы (превышающей 20% от максимальной) из-за внутримышечного давления резко ограничивается кровоснабжение, а, следовательно, и обеспечение мышц кислородом.
Образующаяся в процессе анаэробного гликогенолиза (гликолиза) молочная кислота медленно диффундирует в кровь. Поэтому по мере образования она скапливается внутри сокращающихся мышечных клеток, что ведет к сдвигу их активной реакции в кислую сторону. По мере накопления молочной кислоты происходит угнетение вплоть до полного торможения активности ключевых гликолитических ферментов мышечных клеток, в частности фосфорилазы и фосфофруктокиназы. В результате снижается скорость гликолиза (образования молочной кислоты) и количество энергии (АТФ), образующейся в единицу времени за счет лактацидной системы.
Таким образом, скорость образования (концентрация) в сокращающихся мышцах молочной кислоты регулируется по механизму отрицательной обратной связи — торможением — конечным продуктом реакции: чем больше скорость образования (накопления) молочной кислоты, тем сильнее торможение, замедляющее анаэробный гликогенолиз (гликолиз). Поэтому емкость лактацидной системы в основном лимитируется не содержанием ее энергетических субстратов — углеводов (мышечного гликогена и глюкозы крови), а количеством (концентрацией) конечного продукта деятельности этой системы — молочной кислоты.
В этой связи при выполнении, мышечной работы, энергетическое обеспечение которой осуществляется преимущественно лактацидной системой, никогда не происходит резкого истощения гликогена в работающих мышцах, а тем более в печени.
Ёмкость лактацидной системы. Подсчитаем её.
Среднее содержание гликогена в мышцах — около 15 г/кг, или около 80 мМ глюк. ед (молекулярный вес глюкозы = 180). При работах с максимальной для лактацидной системы нагрузкой в результате анаэробного гликогенолиза в рабочих мышцах расходуется, по-видимому, не более 20 мМ глюк. ед/кг. Таким образом, во всей активной мышечной массе (20 кг) расходуется в целом примерно 400 мМ, или 0,4 М глюк. ед. При анаэробном глюкогенолизе 1 М глюк. ед. дает энергию, достаточную для образования 3 М АТФ.
Следовательно, при анаэробном расщеплении 0,4 М глюк. ед. возможен ресинтез 1,2 М АТФ, что эквивалентно общей энергопродукции за счет анаэробного расщепления мышечного гликогена в 12000 ккал. Эти величины и характеризуют максимальную энергетическую емкость лактацидной системы (см. табл. 210210903).
Другой способ подсчета емкости лактацидной системы — по максимальному количеству образующейся в мышцах молочной кислоты.
Максимальная концентрация молочной кислоты в работающих мышцах достигает у нетренированного человека 250 мг/100 г., или 2,5 г/кг. В этом случае во всей массе активных мышц (20 кг) общее содержание молочной кислоты составляет 50 г, а при учете ее молекулярного веса (90) — около 0,6 М. При анаэробном расщеплении одной глюкозной единицы гликогена освобождается энергия, достаточная для ресинтеза 3 молекул АТФ. При этом образуются 2 молекулы молочной кислоты.
Следовательно, при образовании 0,6 М молочной кислоты освобождается энергия, достаточная для ресинтеза — 0,9 М АТФ (около 9 ккал).
В действительности же при работе общее количество образующейся в мышцах молочной кислоты превышает рассчитанное по ее максимальной концентрации, так как молочная кислота непрерывно диффундирует из мышечных клеток в кровь. Следовательно, и емкость лактацидной системы должна превышать те величины, которые рассчитываются по максимальной концентрации лактата. Энергетическая емкость лактацидной системы примерно в 2,5 раза больше, чем фосфагенной системы (табл. 210210903).
Кислородная (окислительная) энергетическая система
При непрерывном поступлении кислорода в митохондрии мышечных клеток действует кислородная система энергопродукции (ресинтеза АТФ). При работах аэробного характера с повышением интенсивности (мощности) выполняемой нагрузки увеличивается количество O2, потребляемого мышцами (и всем организмом в целом) в единицу времени.
Поскольку между скоростью потребления O2 (л/мин) и мощностью работы (Вт) аэробного характера существует прямо пропорциональная (линейная) зависимость, интенсивность аэробной работы можно характеризовать скоростью потребления кислорода (рис[V.G.44].). При определенной, индивидуальной для каждого человека нагрузке достигается максимально возможная для него скорость потребления O2 — максимальное потребление кислорода (МПК). Для физиологической оценки относительной мощности выполняемой человеком аэробной мышечной работы её оценивают по относительной скорости потребления O2, т.е. выраженному в процентах отношению скорости потребления O2 (л/мин) при выполнении данной работы к МПК (% МПК).
Энергетические субстраты. Для энергетического обеспечения мышечной работы кислородная система может использовать в качестве субстратов окисления («горючего») все основные питательные вещества — углеводы (гликоген и глюкозу), жиры (жирные кислоты) и белки (аминокислоты).
Вклад белков (аминокислот) в общую аэробную энергопродукцию мышц весьма мал и потому может практически не учитываться. До тех пор, пока углеводы и жиры доступны мышце, именно они используются как субстраты аэробной (окислительной) энергопродукции для поддержания мышечных сокращений.
Соотношение между окисляемыми углеводами и жирами определяется относительной мощностью аэробной работы (% МПК): чем больше относительная мощность выполняемой работы, тем относительно выше энергетический вклад окисляемых углеводов и соответственно меньше вклад окисляемых жиров в общую энергопродукцию работающих мышц (рис[V.G.45].).
Во время выполнения легкой работы при потреблении O2 до 50 % МПК (с предельной продолжительностью до нескольких часов) большая часть энергии для сокращающихся мышц образуется за счет окисления жиров. Во время более тяжелой работы (при потреблении O2 более 60 % МПК) значительную часть энергопродукции обеспечивают углеводы. При работах, близких к МПК, подавляющая часть аэробной энергопродукции идет за счет окисления углеводов.
Таким образом, при работе очень большой мощности основными энергетическими субстратами в работающих мышцах служат углеводы. Они расщепляются главным образом аэробно (окисляются) при работе продолжительностью до нескольких десятков минут и в значительной мере анаэробно (гликолитически) при менее продолжительной работе.
Аэробное расщепление углеводов (гликогена и глюкозы) вплоть до стадии образования пировиноградной кислоты идет по тому же пути, что и при анаэробном гликолизе. В последнем случае из-за недостатка O2 (или уменьшения отношения НАД / НАДН+) пировиноградная кислота превращается (восстанавливается) в молочную кислоту при участии фермента лактатдегидрогеназы (ЛДГ). В аэробных условиях пировиноградная кислота не восстанавливается до молочной кислоты, а окисляется. При этом образуются конечные продукты окисления — CO2 и H2O. Суммарное уравнение для аэробного расщепления (окисления) глюкозы (C6H12O6) выглядит следующим образом:
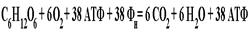
Из этого уравнения следует, что при полном аэробном расщеплении одной молекулы глюкозы образуются 38 молекул АТФ, из которых 2 молекулы АТФ образуются на гликолитическом этапе расщепления глюкозы до пировиноградной кислоты, а 36 молекул АТФ — за счет кислородной системы. На этапах анаэробного расщепления из одной глюкозной единицы мышечного гликогена образуются 3 молекулы АТФ. Поэтому при полном окислении одной глюкозной единицы гликогена образуются 39 молекул АТФ (3 + 36). Следовательно, аэробное расщепление глюкозы дает в 19 раз больше продукции АТФ, чем анаэробный гликолиз (38 молекул АТФ против 2 молекул АТФ).
Окисление глюкозной единицы из мышечного гликогена обеспечивает образование в 13 раз большего числа молекул АТФ, чем при анаэробном гликогенолизе (39 молекул АТФ против 3 молекул АТФ).
Таким образом, с точки зрения расходования горючего (углеводов) кислородная система значительно более эффективна, чем анаэробная лактацидная: при одинаковом расходовании углеводов за счет их окислительного расщепления ресинтезируется во много раз больше АТФ, чем при анаэробном гликогенолизе (гликолизе).
Приведенное выше уравнение позволяет также вычислить кислородный эквивалент АТФ при окислении углеводов, т.е. количество O2, необходимое для окислительного ресинтеза 1 моля АТФ.
Как следует из уравнения, для образования 1 моля АТФ при аэробном расщеплении глюкозы требуется 0,158 молей O2 (6: 38). 1 моль O2, равен 22,4 л O2, следовательно, на образование 1 моля АТФ требуется 3,54л O2 (0,158х22,4). Для образования же 1 моля АТФ при аэробном расщеплении гликогена требуется несколько меньшее количество кислорода: (6: 39) × 22,4 = 3,45 л O2. Соответственно при окислении гликогена расходование 1 моля O2 ведет к образованию 6,5 моля АТФ (39: 6), а при окислении глюкозы — 6,3 моля АТФ (38: 6).
Таким образом, с точки зрения энергетической эффективности O2 мышечный гликоген — более выгодный субстрат для аэробной энергопродукции по сравнению с глюкозой, которую мышцы получают из крови.
Мышечный гликоген является предпочтительным субстратом окисления во время интенсивной мышечной работы. Скорость его расходования находится в прямой связи с относительной мощностью работы (% МПК) и в обратной связи с содержанием в мышце. Чем больше мощность работы (сила сокращений мышц), тем выше скорость расходования мышечного гликогена (см. рис[V.G.46].). До мощности работы примерно 70 % МПК гликоген подвергается главным образом (если не исключительно) аэробному гликогенолизу. При более высоких нагрузках резко увеличивается скорость анаэробного гликогенолиза. При анаэробном гликогенолизе ресинтезируется в 13 раз меньше АТФ, чем при аэробном расщеплении гликогена. Поэтому для образования одинакового количества АТФ скорость расходования гликогена при анаэробном расщеплении (лактацидной системой) должна в 13 раз превышать скорость расходования гликогена окислительной системой.
Это объясняет резкое (экспоненциальное) повышение скорости расходования гликогена при увеличении мощности работы сверх 70% МПК.
По мере уменьшения содержания мышечного гликогена в процессе работы постоянной мощности скорость его расходования снижается (см. рис. 23, Б), а использование работающими мышцами глюкозы из крови увеличивается. Скорость утилизации глюкозы из крови работающими мышцами также растет с увеличением мощности работы (в дополнение к повышению скорости расходования мышечного гликогена). Таким образом, соотношение между расходованием гликогена и глюкозы зависит как от мощности, так и от продолжительности работы. Например, в начале работы на уровне 70% МПК (длительностью 1,5—2 ч) потребление глюкозы мышцами составляет лишь 10—15% от общего расхода углеводов, а в конце работы — до 50% и более.
При расчете ёмкости кислородной системы, обеспечиваемой углеводами, следует учитывать запас гликогена в мышцах и печени, а также возможность печени в процессе работы образовывать глюкозу не только за счет расщепления гликогена (гликогенолиза), но и за счет образования глюкозы (глюконеогенеза) из лактата и других веществ (аминокислот, пирувата, глицерола), попадающих в печень с кровью.
Если принять среднюю концентрацию гликогена в мышцах равной 15 г/кг, то общее его содержание в 20 кг активной мышечной массы равно 300 г (или около 1,7 М глюк. ед). Степень расходования гликогена разными мышцами неодинакова, так как зависит от степени участия каждой из мышц в выполняемой работе. В целом можно допустить, что во всех 20 кг активных мышц содержание гликогена может уменьшиться максимально на 80%. Таким образом, максимальное количество расходуемого гликогена вряд ли превышает 250 г (около 1,5 М глюк. ед.). При окислении этого количества гликогена кислородная система продуцирует энергию, достаточную для образования примерно 60 М АТФ (1,5×39).
У мужчин весом около 70 кг общий запас гликогена в печени равен примерно 70—80 г. Во время работы его расходуется не более 60 г. Если даже принять, что вся глюкоза, образующаяся из этого количества гликогена, используется мышцами, то она обеспечивает дополнительно ресинтез еще около 15 М АТФ. К последней величине следует прибавить некоторое количество глюкозы, образующейся в печени за счет глюконеогенеза. Эта величина, по-видимому, составляет не более 10—20% от общей продукции глюкозы печенью при кратковременной тяжелой и до 45% при длительной работе. Таким образом, общее количество АТФ, которое могут продуцировать рабочие мышцы за счет потребления глюкозы из крови, вряд ли превышает 20 М.
Приведенные расчеты позволяют заключить, что емкость кислородной системы, использующей в качестве субстрата окисления углеводы, составляет величину порядка 80 М АТФ, или 800 ккал. Только за счет окисления доступных запасов углеводов нетренированный человек может, например, пробежать дистанцию в 15 км.
Другим важнейшим субстратом кислородной системы служат жиры (липиды). Наибольшая часть жиров в теле человека находится в форме триглицеридов. Молекула триглицерида состоит из четырех частей — одной молекулы глицерида (глицерола) и трех молекул жирных кислот. Образование АТФ происходит в основном за счет окисления молекул жирных кислот, которые освобождаются при расщеплении триглицерида. Поэтому чаще всего говорят об образовании АТФ за счет окисления жирных кислот. Этот процесс происходит только в митохондриях и называется бета-окислением.
В среднем 1 моль смеси жирных кислот, характерных для человека, обеспечивает образование 138 молей АТФ. Таким образом, при аэробном расщеплении молекулы жирной кислоты образуется в несколько раз больше молекул АТФ, чем при аэробном распаде молекулы глюкозы (или глюкозной единицы из гликогена).
При аэробном использовании 1 моля глюкозы образуется 38 молей АТФ, т. е. 1 г окисленной глюкозы обеспечивает ресинтез 0,21 М АТФ. Это соответствует 2,1 ккал. При полном сгорании 1 г углеводов в калорической бомбе выделяется, как известно, 4,1 ккал. В мышце 2,1 ккал идет на ресинтез фосфагенов, а остальные 2 ккал (около 50 % энергии) выделяются в виде тепла. То же справедливо и в отношении жиров, полная энергия 1 г которых равна 9,1 ккал, однако для ресинтеза АТФ используются лишь 5 ккал.
При окислении 1 моля пальмитиновой кислоты (молекулярный вес = 256) образуется 128 молей АТФ, т.е. каждый грамм этой жирной кислоты при полном окислении обеспечивает ресинтез 0,5 моля АТФ, что эквивалентно примерно 5 ккал. Иначе говоря, при одинаковом по весу расходе углеводов и жиров последние могут обеспечить ресинтез в два с лишним раза большего количества АТФ, чем при окислении углеводов.
Жиры обладают наибольшей энергетической емкостью из всех остальных мышечных источников энергии. 1 моль АТФ дает около 10 ккал, 1 моль КФ — около 10,5 ккал, 1 моль глюкозы при анаэробном расщеплении — около 50 ккал, при аэробном расщеплении (окислении) — около 700 ккал, а 1 моль жиров при окислении обеспечивает 2400 ккал.
Необходимо отметить, что запасы жиров в теле человека велики — жиры составляют от 10 до 30 % его веса. Только небольшая часть жиров (до 5 % всех жировых запасов) находится в мышцах в виде триглицеридов, концентрация которых в мышцах составляет от 5 до 15 г/кг. Основная масса жира находится в жировых депо тела. Отсюда освободившиеся при расщеплении (липолизе) свободные жирные кислоты попадают в ток крови, а затем захватываются из крови мышцами.
Во время работы мышцы используют как свободные жирные кислоты из крови, так и собственные освобождающиеся из триглицеридов. При работе на уровне 50 – 70 % МПК вклад этих двух источников примерно одинаков. Приблизительные подсчеты показывают, что за счет окисления всех запасенных в теле жиров активная мышечная масса в 20 кг может ресинтезировать несколько тысяч молей АТФ (см. табл. 210210903). Эта величина характеризует огромную энергетическую емкость кислородной системы, использующей жиры в качестве субстрата окисления. Запасов жиров вполне достаточно для энергетического обеспечения непрерывной ходьбы в течение 7—10 дней.
Кислородная система, использующая в качестве субстратов окисления как углеводы, так и жиры, обладает наибольшей энергетической емкостью из всех других энергетических систем. Ее емкость во много тысяч (!) раз превышает емкость лактацидной и фосфагенной систем (см. табл. 210210903). Поэтому кислородная энергетическая система обеспечивает возможность выполнения работы большой продолжительности — от нескольких минут до многих часов и даже дней.
Приведенные выше данные позволяют сопоставить энергетические характеристики углеводов и жиров как субстратов окисления (табл. 3). Жиры являются самым легким «горючим», если сравнивать энергию, содержащуюся в единице веса вещества (удельная энергетическая емкость). Для образования одинакового количества АТФ расходуется по весу почти в 2,5 раза меньше жиров, чем глюкозы. В мышце каждый грамм гликогена связан с 2,7 г воды. Поэтому каждому грамму гликогена соответствует 3,7 г комплекса «гликоген + вода». Это снижает энергетическую удельную емкость мышечного гликогена до 0,07 моль АТФ/г комплекса «гликоген + вода». В действительности гликоген — самое тяжелое «горючее». Жиры имеют также преимущество перед углеводами по общей энергетической емкости. Все это делает их пригодными для выполнения длительной работы относительно небольшой мощности.
Однако углеводы имеют серьезное преимущество перед жирами, когда сравнивается количество АТФ, образующееся на единицу потребляемого O2, (энергетическая эффективность O2). При таком сравнении углеводы на 10—13% эффективнее, чем жиры. Если выполняется очень интенсивная аэробная работа (близкая к МПК, т. е. околомаксимальная аэробная работа), она в большой степени лимитируется скоростью потребления O2. В этом случае углеводы как субстрат окисления имеют неоспоримое преимущество перед жирами, так как для образования одинакового количества энергии (АТФ) при окислении углеводов затрачивается меньшее количество O2, чем при окислении жиров. Особенно эффективно в этом случае окисление мышечного гликогена, имеющего наибольшую энергетическую эффективность O2.
Таблица 210231404
Энергетические характеристики окислительного использования углеводов и жиров
| Показатели | Мышечный гликоген (глюк.ед.) | Глюкоза | Жирные кислоты |
| Удельная энергетическая емкость: | |||
| моль АТФ/моль субстрата | |||
| моль АТФ/г субстрата | 0,22 | 0,22 | 0,50 |
| Общая энергетическая емкость (на 20 кг мышечной массы), моль АТФ | |||
| Энергетическая эффективность O2, моль АТФ/моль O2 | 6,50 | 6,34 | 5,61 |
| Кислородный эквивалент АТФ, л O2 / моль АТФ | 3,45 | 3,54 | 4,05 |
| Максимальная мощность (на 20 кг активной мышечной массы), моль АТФ/моль субстрата · с-1 | 0,8 | 0,5 | 0,4 |
Наконец, общее количество энергии (АТФ), которое может продуцироваться в единицу времени за счет окисления углеводов (особенно мышечного гликогена), вдвое больше, чем при окислении жиров. Такая относительно сниженная мощность энергопродукции при окислении жиров по сравнению с окислением углеводов позволяет понять, почему участие жиров в энергообеспечении уменьшается по мере повышения мощности выполняемой работы.
[a] Мышечное волокно
[b] Структурные компоненты миона: эндомизий (коллагеновые волокна, фиброциты, гемокапилляр, моторная бляшка) + мышечное волокно (базальная мембрана, миосателлитоцит, миосимпласт).
[c] Синонимы – нейро‑моторная единица, двигательная единица.
[d] Выраженность этой способности у различных атипичных кардиомиоцитов существенно разнится. У рабочих кардиомиоцитов практически нет.
[e] Расположены в капсуле, в составе нервно-мышечного веретена fusus neuromuscularis
[f] Синоним - фазические
[g] Под световым микроскопом мышечное веретено представляет собой вытянутую структуру, расширенную посередине за счет капсулы и напоминающую по форме веретено, что и обусловило ее название.
[h] Тонус [лат. tonus < гр. tonos напряжение] — состояние длительного возбуждения нервной или мышечной ткани, не сопровождающееся развитием утомления.
[i] Подробнее читайте в Приложение 709261417. Энергетика мышечного сокращения.
[j] Синоним – окислительные.
[k] Huxley H.E., Hanson J.
[l] Синоним – электромеханический каплинг
[m] Потенциалуправляемых (электроуправляемых)
[n] Также у нейронов
[o] инозитолтрисфосфатом
[p] Разность между запросом и реальным потреблением кислорода в этот период называется кислородным дефицитом (см. 19.3).
[1] Необходимо знать ХОРОШО! С материалом этого раздела Вы должны быть уже знакомы.
[2] Необходимо знать ХОРОШО! Но с материалом этого раздела Вы должны быть уже хорошо знакомы. Подробности в учебнике.
[3] Необходимо знать ХОРОШО! Но с материалом этого раздела Вы должны быть уже хорошо знакомы. Подробности в учебнике.
[4] Необходимо знать ХОРОШО! Но с материалом этого раздела Вы должны быть уже хорошо знакомы.
[5] Необходимо знать ОЧЕНЬ ОЧЕНЬ ХОРОШО, в том числе и двоечникам! Но с материалом этого раздела Вы должны быть уже хорошо знакомы.
[6] Это Вы должны уже знать. Если забыли, вспомните. Воспроизводить могут только отличники и хорошисты.
[7] Необходимо знать ХОРОШО! Но с материалом этого раздела Вы должны быть уже хорошо знакомы.
[8] Необходимо знать ХОРОШО!
[9] Необходимо знать ХОРОШО! Подробнее мы будем знакомиться с этим типом мышечных волокон при изучении сенсорных систем.
[10] Необходимо знать ХОРОШО!
[11] Для ознакомления! Подробно в учебнике.
[12] Необходимо знать ХОРОШО!
[13] Необходимо знать ОЧЕНЬ ХОРОШО!
[14] Необходимо знать ОЧЕНЬ ОЧЕНЬ ХОРОШО, в том числе и двоечникам!
[15] Необходимо знать ОЧЕНЬ ОЧЕНЬ ХОРОШО, в том числе и двоечникам!
[16] Необходимо знать ОЧЕНЬ ОЧЕНЬ ХОРОШО, в том числе и двоечникам!
[V.G.1](см. рис. 21)
[V.G.2](см. также рис. 21, е)
[Мф3]++501+C.74
[Мф4]++501+C.74
[Мф5]++501+C.74
[V.G.6](см. рис. 22, б)
[V.G.7]210211139
[V.G.8]210211146
[V.G.9]210211148
[Мф10]++760+ С.57
[Мф11]++597+ С.294
[Мф12]Вопрос 2.
[Мф13]++597+ С.294
[Мф14]
[Мф15]++597+ С.292
[Мф16]I Сам придумал
[Мф17]в презентации лучшего качества
[Мф18]++51+ С.693
[Мф19]Камкин С.329
[Мф20]++736+ С.125
[Мф21]++736+ С.125
[Мф22]Камкин С.329
[Мф23]Современная теория мышечного сокращения и расслабления.
[Мф24]++501+С.74; --147-С.23
[Мф25]--147-С.23
[Мф26]++601+С.76
[Мф27]++501+С.74
[Мф28]Распространение электрической волны внутрь волокна приводит, в свою очередь, к деполяризации мембран цистерн продольных трубочек саркоплазматического ретикулума (Коц).
[Мф29]++597+С.293
[Мф30]++501+С.72
[Мф31]++810+ С.101 Орлов С.
[Мф32]++810+ С.101 Орлов С.
[Мф33]++810+ С.101 Орлов С.
[Мф34]++810+ С.101 Орлов С.
[Мф35]++810+ С.101 Орлов С.
[Мф36]++597+ С.294 №7
[V.G.37](см. 6.5.2)
[Мф38]++597+ С.294 №8
[Мф39]++736+С.100
[Мф40]++736+С.102
[V.G.41]при расщеплении 1 М КрФ (М-1 моля)???
[Мф42]++736+С.101
[V.G.43]23
[V.G.44]24
[V.G.45]25
[V.G.46]23
Date: 2015-09-19; view: 826; Нарушение авторских прав