Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Электроэнцефалографические исследования восприятия
|
|
Изучение физиологических основ восприятия проводится с помощью макропотенциалов мозга: фоновой электроэнцефалограммы, а также вызванных и событийно-связанных потенциалов, причем использование ЭЭГ имеет два разных аспекта. Первый связан с выявлением параметров ЭЭГ, имеющих отношение к кодированию стимула. Второй, более традиционный, представляет собой изучение параметров ЭЭГ отдельных зон коры или показателей их совместного взаимодействия при восприятии стимулов.
ЭЭГ-показатели восприятия. Среди ритмов ЭЭГ наибольшее J внимание исследователей в этом плане привлекает альфа-ритм (8 —12 Гц), который регистрируется преимущественно в задних отделах коры в состоянии спокойного бодрствования. Известно, что при предъявлении стимулов имеет место подавление или «блокада» альфа-ритма: причем она тянется тем больше, чем сложнее изображение.
Существует немало предположений, касающихся роли альфа-ритма в обеспечении активности мозга как целого. Основоположник кибернетики Н. Винер и вслед за ним ряд других исследователей считали, что этот ритм отражает механизм временного сканирования («считывания») информации и тесно связан с механизмами восприятия и памяти. Действительно по некоторым данным биопотенциальное поле альфа-ритма связано с функцией сканирования информации в реальном времени и тем самым связано с процессами мечжмодальной сенсорной интеграции в процессах восприятия и памяти. По другим представлениям, альфа-ритм связан с действием селектирующих механизмов мозга, выполняющих функцию резонансного фильтра, и таким образом регулирующих поток сенсорных импульсов.
Поиск электрофизиологических показателей восприятия в па-
162 • ■;.'■, •'. ■**-
раметрах альфа-ритма опирается на представление о том, что воспринимаемая человеком информация кодируется комбинациями фаз и частот периодических нейронных процессов, которые находят свое отражение в характеристиках альфа-ритма. Предполагается, что такая особенность ЭЭГ, как пакет волн, создается синхронизированной когерентной активностью группы нейронов, расположенных в разных участках мозга и образующих ансамбль. Предположительно все волны одного пакета хранят информацию об одном образе или его части и при восприятии опознается только тот образ, который закодирован ритмической активностью наибольшего числа нейронов в каждый данный момент времени (Лебедев, 1985).
Экспериментально было показано, что значения параметров колебаний системы нейронов, оцениваемые, в частности, по расстоянию между соседними спектральными пиками внутри альфа-диапазона, могут служить аргументами уравнений, предсказывающих некоторые особенности восприятия. Например, чем больше период доминирующих колебаний в ЭЭГ человека и чем больше разнообразие воспринимаемых и ожидаемых стимулов, тем медленнее осуществляется их восприятие.
Наряду с этим существуют исследования пространственно-временных отношений потенциалов мозга при восприятии сенсорной информации. В отличие от предыдущего этот подход учитывает два фактора: время восприятия и его мозговую организацию. Речь идет о множественной регистрации ЭЭГ из разных зон коры больших полушарий в процессе восприятия. Поскольку нейронные ансамбли, участвующие в переработке информации, распределены по разным отделам мозга, в первую очередь, коры больших полушарий, логично считать, что перцептивный акт будет сопровождаться изменением пространственного соотношения ЭЭГ. Действительно, изучение дистантной синхронизации биопотенциалов коры (Ливанов, 1977) в ходе зрительного восприятия позволяет выявить следующее: вначале наблюдается преимущественная активация задних отделов коры обоих полушарий, затем в процесс вовлекаются передние отделы правого полушария. Узнавание связано с активным включением в процесс центральных и фронтальных зон коры.
Изменения биоэлектрической активности мозга в процессе перцептивного акта характерны для всех видов сенсорных модально-
<•• -. 163
ill
 стей. Однако в зависимости от вида анализатора эти изменения имеют разную пространственную картину. Как правило, на ранних этапах перцепции наибольшую активность демонстрируют проекционные зоны (зрительная, слуховая, соматосенсорная), а на завершающих этапах в процесс включаются передние отделы коры.
стей. Однако в зависимости от вида анализатора эти изменения имеют разную пространственную картину. Как правило, на ранних этапах перцепции наибольшую активность демонстрируют проекционные зоны (зрительная, слуховая, соматосенсорная), а на завершающих этапах в процесс включаются передние отделы коры.
Вызванный потенциал как коррелят перцептивного акта. На-1 иболее адекватным инструментом изучения физиологической природы перцептивного процесса является метод регистрации вызванных (ВП) или событийно-связанных потенциалов. Последнее обусловлено тем, что во-первых, ВП — это реакция мозга на внешнее раздражение, по длительности сопоставимая со временем перцептивного акта, во-вторых, ВП, взятые отдельно в каждой зоне коры, имеют свои характеристики, обусловленные функциональными особенностями этой зоны. Таким образом, при предъявлении стимула любой модальности (зрительного, звукового, тактильного и др.) в разных зонах коры возникают специализированные реакции, сопоставимые по | длительности с реальным психологическим процессом восприятия.
Первые исследования в этом направлении ставили своей целью I соотнести конфигурацию и параметры ВП с типом стимула и вы-1 делить варианты ответов, связанных с содержанием и физическими характеристиками стимула, например, установить отвечает ли мозг специфическим ответом на геометрические фигуры (круг, квадрат) или определенные слова. Предпринимались даже попытки определять по конфигурации ВП, какой стимул предъявлялся испытуемому. Несмотря на определенные положительные результаты, эти исследования не получили развития.
На первый план выступил иной подход. В его основе лежали представления о том, что компонентная структура ВП отражает не конкретное содержание или признак воспринимаемого стиму-; ла, а некоторые общие последовательные операции головного моз- \ га по обработке стимула.
ВП и последовательная модель переработки информации. Бы- j ло установлено, что в конфигурации ВП можно выделять компоненты двух типов: ранние специфические (экзогенные) и поздние неспецифические (эндогенные) компоненты. Обработка физических параметров стимула (сенсорный анализ) связывается с деятельностью специфических сенсорных систем и находит свое от-
ражение преимущественно в параметрах экзогенных компонентов ВП. В зрительной модальности эти компоненты регистрируются в интервале до 100 мс от начала ответа, в слуховой и соматосенсор-ной модальностях — еще раньше, в пределах 10 — 50 мс от начала стимуляции. Эндогенные компоненты ВП, возникающие позднее и длящиеся до окончания ответа, предположительно отражают этапы более сложной обработки стимула: формирование образа, сличение его с эталонами памяти, принятие перцептивного решения. При изучении структуры перцептивного акта с помощью вызванных потенциалов так же, как и при изучении внимания, получают разностные кривые, «в чистом виде» представляющие когнитивную составляющую биоэлектрической активности (рис. 5.2).
| N1 |
 5 мкВ/дел.
5 мкВ/дел.
75 мс/дел.
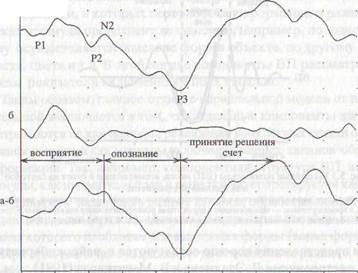
Рис. 5.2 Выделение когнитивной составляющей ответа.
а. Ответ в условиях опознания значимого стимула у здорового испытуемого (45 лет): P1-N1-P2 собственно
ответ на стимул, N2 —170 мс и РЗ — 357 мс.
б. Ответ на тот же стимул без его опознания;
а-б. Разность между ответами в условиях распознавания и в стандартных условиях без распознавания. Когнитивный комплекс. Восприятие начинается на 76 мс, опознание и дифференцировка — компонент N2 — 170 мс, принятие решения и запоминание — компонент РЗ — 354 мс (по В.В.Гнездицкому, 1997).
164
165

 Интерпретация процессов, отражающихся в параметрах и конфигурации ВП, может осуществляться в контексте разных моделей: последовательной, параллельной и циклической.
Интерпретация процессов, отражающихся в параметрах и конфигурации ВП, может осуществляться в контексте разных моделей: последовательной, параллельной и циклической.
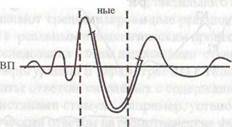
Первую иллюстрирует концепция «информационного синтеза» A.M. Иваницкого (1986). Согласно этой концепции ранние компоненты ВП в интервале 0 — 100 мс связаны с оценкой физических параметров стимула, в среднелатентных компонентах в интервале 100 — 200 мс находят отражение процессы синтеза текущей сенсорной информации с содержанием памяти, а в поздних компонентах ВП отражается когнитивная оценка стимула как такового (рис. 5.3).
| значимость \ Щ |

 чувствительность
чувствительность
| стимул |
 ранние | проме- | поздние I жуточ-1
ранние | проме- | поздние I жуточ-1
время, мс
Рис. 5.3 Вызванные потенциалы (ВП) мозга, возникающие в ответ на световой
или кожный раздражитель.
Видно, что ранние волны соотносятся с чувствительностью, поздние — с мотивацией, значимостью стимула, а промежуточные — одновременно с двумя этими психофизическими показателями. Стрелкой на временной шкале обозначен момент предъявления стимула (по А.М.Иваницкому, 1999).

 Эти представления хорошо согласуются с нейрокибернетиче-скими построениями Д. Эделмена и В. Маунткасла (1981), согласно которым повторный возврат возбуждений является мозговой основой всех психяческих процессов. Так, например, момент возникновения образа совпадает по времени со встречей в коре двух информационных потоков, один из которых представляет собой внешний сигнал, а другой память. Предполагается, что эта память и есть возникающая в сознании человека частица «я», по отношению к которой, сигнал воспринимается как нечто внешнее (см. главу 10).
Эти представления хорошо согласуются с нейрокибернетиче-скими построениями Д. Эделмена и В. Маунткасла (1981), согласно которым повторный возврат возбуждений является мозговой основой всех психяческих процессов. Так, например, момент возникновения образа совпадает по времени со встречей в коре двух информационных потоков, один из которых представляет собой внешний сигнал, а другой память. Предполагается, что эта память и есть возникающая в сознании человека частица «я», по отношению к которой, сигнал воспринимается как нечто внешнее (см. главу 10).
166
Изложенные выше представления объяснимы с помощью последовательной модели переработки информации, согласно которой каждая стадия перцептивного процесса наступает не ранее, чем реализуется предыдущая.
ВП и другие модели переработки информации. В то же время имеются данные, трудно объяснимые в контексте последовательной модели переработки информации. Например, при регистрации зрительных ВП на особо сложные стимулы параметры этих стимулов могут обрабатываться во всем временном диапазоне ВП. Последнее предполагает наличие иных моделей переработки информации: параллельной и циклической. Причем в рамках обоих моделей ВП имеют различное толкование.
В основе параллельной модели лежит представление о существовании в зрительной системе нескольких параллельно работающих подсистем, в которых переработка информации о разных признаках стимула происходит независимо, например, по одному каналу осуществляется описание формы объекта, по другому — контраста, цвета и т.д. В этой логике компоненты ВП рассматриваются как показатели включения разных каналов.
Таким образом, главное отличие параллельной модели от последовательной заключается в том, что отдельные компоненты здесь рассматриваются не как маркеры реализации конкретных стадий обработки информации, а как маркеры включения разных каналов обработки информации. Так, например, компонентная структура ВП на такие стимулы, как шахматные поля и решетки интерпретируется как последовательность включения разных каналов, обрабатывающих информацию о яркости (яркостный канал) и о пространственном контрасте, анализ которого необходим для выделения формы (канал формы).
При этом различия в выраженности относительно ранних и поздних компонентов ВП в ответах на низкие и высокие пространственные частоты отражают не временную последовательность процессов обработки высоких и низких пространственных частот, а различную пространственную локализацию генераторов ВП на разные пространственные частоты (Зислина, 1978). В принципе эти предположения имеют определенные основания, поскольку опираются на электрофизиологические данные, касающиеся источников генерации ВП, а те подтверждают, что относительно ранние компоненты ВП на шахматные
.. 167

|

|
I
поля и решетки с латентным периодом 75 —125 мс генерируются в стриарной коре, а более поздние имеют своим источником парастри-арную область.
Циклическая модель предполагает, что в перцептивном процессе могут существовать отдельные и независимые перцептивные циклы с разной внутренней структурой и продолжительностью отдельных стадий. При циклической обработке информации, по-видимому, происходит повторное активирование одних и тех же нейронных популяций. Переработка информации при этом обеспечивается не только однократными специфическими реакциями нейронов, но и их повторным вовлечением в фазический процесс.
При трудной перцептивной задаче правильное опознание с большей вероятностью происходит на поздних этапах переработки информации. По мере привыкания возрастает вероятность опознания сигнала на более ранних фазах переработки. При этом предполагается, что число компонентов ВП определяется количеством фаз переработки информации в корково-подкорковых нейронных сетях (Переслени с соавт., 1987). Таким образом, в одном и том же интервале времени после предъявления одних и тех же стимулов могут возникать различающиеся по числу компонентов и конфигурации ВП. И наоборот, внешне одни и те же компоненты ВП могут соответствовать разным стадиям обработки стимула.
Эти представления ставят под сомнение возможность существования фиксированной схемы обработки информации по типу «постоянный стимул — неизменная реакция». В то же время логически непротиворечивое обоснование вышеописанные факты получают с позиций гипотезы когнитивных детерминант организации перцептивного акта. Согласно этой гипотезе порядок следования и набор операций (и соответствующих компонентов ВП) в каждом перцептивном акте непостоянен (от пробы к пробе) и определяется в процессе поступления информации и по ходу ее обработки. Решающую роль при этом будут играть условия задачи и опыт субъекта в организации информации.
Date: 2015-08-15; view: 420; Нарушение авторских прав