Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Антигеннезависимая дифференцировка Т-лимфоцитов
|
|
 Родоначальной клеткой Т-лимфоцитов, как и всех клеток крови, является полипотентная стволовая гемопоэтическая клетка. Её маркером является CD 34.
Родоначальной клеткой Т-лимфоцитов, как и всех клеток крови, является полипотентная стволовая гемопоэтическая клетка. Её маркером является CD 34.
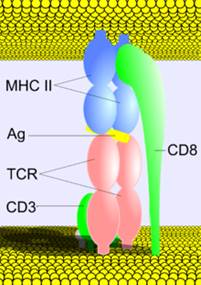
Ранние предшественники Т-лимфоцитов мигрируют из костного мозга в тимус, где происходит антигеннезависимая дифференцировка Т-клеток под влиянием «клеток нянек», эпителиальных клеток тимуса, а так же гормонов тимуса (α- и β-тимозины, тимулин /сывороточный фактор тимуса/, тимопоэтин, тимический гуморальный фактор). Самыми ранними маркерами тимоцитов являются CD7, CD2. В тимусе Т-лимфоциты дифференцируются в иммунокомпетентные клетки и приобретают важную способность к распознаванию антигена. На их наружной мембране появляется (экспрессируется) особый рецептор - Т-клеточный рецептор (ТКР, англ. - TcR, T-cell receptor) для антигена. Причем для каждого антигена (эпитопа) в организме предназначен отдельный лимфоцит или его клональные дочерние лимфоциты-потомки, которые имеют специфичный антигену TcR. Тимоциты одновременно с TcR в процессе дифференцировки приобретают CD3, который тесно связан с Т-клеточным рецептором. CD3 необходим для передачи сигнала от ТКР в цитоплазму. На поверхности тимоцитов появляются также молекулы CD8 и CD4. Это двойные позитивные клетки, т.е. их фенотип (ТКР+, CD3+, CD4+, CD8+) и они α β Место связывания антигена являются молодыми тимоцитами. По своему строению молекулы TcR (ТКР) напоминают иммуноглобулины (Fab-фрагмент) и состоят и альфа- и бета- цепей (TcR αβ их подавляющее большинство) или гамма- и дельта- цепей (TcR γδ). αβ- и γδ – формы TcR весьма сходны по структуре. Каждая цепь ТКР состоит из двух областей (доменов): наружный вариабельный (V), второй – константный (С). Отдельные гены кодирующие всю вариабельную область (V) α и β цепей TcR отсутствуют. Фрагменты вариабельных доменов кодируются тремя группами генов обозначаемых V, D, J. В клеточном геноме гены, кодирующие V-, J- и D-сегменты вариабельной области, представлены в виде многочисленных вариантов. Именно различные сочетания V-, J- и D-сегментов V области, образующиеся в процессе генной перестройки, называемой реаранжировкой, обеспечивают разнообразие молекул ТКР. Таким образом, ограниченное число генов (около 400) может кодировать рецепторы для почти бесконечного числа антигенов (многих миллионов). Причем различные комбинации генов V, D, J –сегментов - это только один из способов достижения многообразия антигенных рецепторов Т-лимфоцитов. Основная функция зрелых Т-лимфоцитов – распознавание чужеродных антигенных пептидов в комплексе с собственными антигенами главного комплекса гистосовместимости (ГКГС) на поверхности антигенпрезентирующих клеток или на поверхности любых клеток-мишеней организма. Для выполнения этой функции Т-лимфоциты должны быть способны распознавать собственные антигены ГКГС. В то же время, Т- клетки не должны распознавать аутоантигены самого организма, связанные с собственными антигенами ГКГС. В связи с этим в тимусе молодые тимоциты проходят селекцию («отбор»), TcR которых соответствует вышеуказанным условиям. Суть позитивной и негативной селекции состоит в следующем:
Позитивная селекция. Т-лимфоциты, ТКР которых обладает способностью распознавать HLA (молекулы ГКГС) стромальных клеток тимуса, выживают, а если нет – то гибнут путем апоптоза. Позитивная селекция – поддержка избирательной выживаемости. Таким образом, выживают только лимфоциты способные распознавать собственные HLA! И эта способность в последующем является важной в функционировании Т-клеток. Кроме этого в тимусе погибают путем апоптоза аутореактивные лимфоциты (лимфоциты имеющие ТКР к антигенным детерминантам собственных тканей). Важно, что при контакте с эпителиоидными клетками тимуса Т-лимфоциты, реагирующие на «своё», разрушаются путем запуска апоптоза (запрограммированной клеточной смерти при активации через CD95 – Fas рецептор). Это отрицательная селекция. В итоге, исчезают аутореактивные клоны клеток и возникает толерантность (неотвечаемость) к «своему». В тимусе около 95 – 97% лимфоцитов погибают в результате процесса селекции. В последующем одна из молекул CD4 или CD8 утрачивается и клетки становятся зрелыми.

Клетки, сохранившие CD4 являются Т-хелперами (Тh) и их ТКР распознает HLA II класса, а сохранившие CD8 – цитотоксическими Т -лимфоцитами и их ТКР обладает способностью распознавать HLA I класса. Из тимуса они мигрируют в периферические лимфоидные органы, где заселяют преимущественно Т-зависимые зоны. На одном Т-лимфоците только один вариант рецептора и только к одномуантигену. TcR прочно связан с CD3.
В частности в лимфоузлах – паракортикальную. Зрелые лимфоциты рециркулируют. Таким образом, АНТИГЕН НЕ ЗАВИСИМАЯ дифференцировка Т-лимфоцитов включает в себя пролиферацию, приобретение специфических маркеров Т-лимфоцитами и образование дифференцированных, зрелых субпопуляций, способных осуществлять характерные для той или иной субпопуляции функции (индукция иммунного ответа, его регуляция, цитотоксичность). В порцессе антигеннезависимой дифференцировки образуются лимфоциты, которые генетически детерминированы к взаимодействию с определенным антигеном и иммунному ответу на этот антиген.
АНТИГЕНЗАВИСИМАЯ ДИФФЕРЕНЦИРОВКА Т-ЛИМФОЦИТОВ
Антигензависмая дифференцировка происходит в периферических органах иммунной системы, если Т-лимфоцит провзаимодействовал с антигеном. Начинается она с момента распознавания антигена и заканчивается образованием клона лимфоцитов, способных _____оказывать специфическое действие как по отношению к антигену, так и к другим иммунокомпетентным клеткам, вступившим во взаимодействие с антигеном. Причем хелперы и цитотоксические лимфоциты распознают антиген по-разному. Так, ХЕЛПЕРЫ (CD4–клетки) распознают АНТИГЕН в комплексе с HLA II КЛАССА, КИЛЛЕРЫ (CD8-клетки) - в комплексе антиген с HLA 1 КЛАССА. Распознавание антигена Т-хелпером является центральным процессом, как в гуморальном иммунном ответе, так и в усилении клеточной формы иммунного ответа.
Специфическими МАРКЕРАМИ для ВСЕЙ ПОПУЛЯЦИИ Т-ЛИМФОЦИТОВ являются имеющиеся на наружной мембране этих клеток антигены CD 3. (Раньше использовался маркер CD 2 - рецептор для эритроцитов барана, что не совсем корректно. Параметры CD антигенов смотри в приложении.)
Маркер Т-лимфоцитов - структура, которая свойственна только Т-лимфоцитам (всем субпопуляциям Т-лимфоцитов) – CD3.
СУБПОПУЛЯЦИИ ЛИМФОЦИТОВ:
Тh лимфоциты. Примерно половина из числа циркулирующих Т-лимфоцитов несут на своей поверхности антиген CD4. Эти Т-лимфоциты функционируют как ХЕЛПЕРЫ, то есть помощники (от англ. To help — помогать), «включающие» популяцию В-лимфоцитов в процесс выработки антител, а Т-эффекторы – в реализацию клеточного иммунитета. Т-хелперы опосредуют свою функцию гуморальными факторами - цитокинами, которые синтезируются этими лимфоцитами в ответ на антигенный стимул. Недостаточность хелперной функции Т-лимфоцитов, наблюдаемая при синдроме приобретенного иммунодефицита (СПИД, одной из важнейших мишеней ВИЧ являются Т-лимфоциты хелперы), приводит к «неотвечаемости» организма на антигенную стимуляцию, что, в конечном итоге, способствует персистенции в организме человека микроорганизмов, развитию злокачественных новообразований и является причиной летального исхода.
Т-хелперы (Th) – стимулируют пролиферацию и дифференцировку как Т-, так и В-лимфоцитов, выделяя цитокины. В зависимости от того, какие цитокины они продуцируют (в зависимости от цитокинового профиля) среди них различают:
Th1 (Т-хелперы первого типа), выделяют ИЛ-2 и γ-интерферон, и в итоге обеспечивают реакции Т-клеточного иммунитета – стимулируют иммунный ответ против внутриклеточных бактерий, противовирусный, противоопухолевый, трансплантационный иммунитет.
Th2 (Т-хелперы второго типа), секретируют ИЛ-4, ИЛ-5, ИЛ-6, ИЛ-10, ИЛ-13 и стимулируют синтез антител, способствуют развитию гуморального иммунного ответа против внеклеточных бактерий, их токсинов, а также образование IgE-антител. Между Th1 и Th2 существует антагонизм: при повышении активности одних, угнетается функция других. В итоге преобладает Т-клеточный (Th1→ T киллеры) или В-клеточный (Th2 →В-лимфоциты →антитела) иммунитет, что во многом зависит от вида антигена. Таким образом, Т-хелперы выполняют хелперно-регуляторную функцию во взаимодействии иммунокомпетентных клеток, направленную на развитие эффекторной фазы иммунного ответа. Именно от Th зависит будет преобладать гуморальный или клеточный иммунный ответ.
 Тk. Среди субпопуляций Т-лимфоцитов различают эффекторные клетки. В связи с тем, что эти эффекторные клетки способны специфически разрушать клетки-мишени, их называют ЦИТОТОКСИЧЕСКИМИ Т-ЛИМФОЦИТАМИ, или Т-КИЛЛЕРАМИ - убийцами (от англ. to kill - убивать). Т-киллер — одна из основных эффекторных клеток клеточноопосредованного иммунитета, которая наряду с другими клетками способна осуществлять лизис клеток мишеней. Роль Т-киллеров очень велика в реализации трансплантационного иммунитета, развитии аутоиммунных заболеваний, в противоопухолевой защите. Тk-лимфоциты (CD8+ клетки) составляют около 20 – 25% от числа циркулирующих Т-лимфоцитов (абсолютноеколичество - 500 – 1200 в 1 мм3 (мкл)), они несут маркерный антиген CD8. Макромолекула CD8 служит корецептором для антигенов главного комплекса гистосовместимости I класса (ГКГС-1). Активированные антигеном цитотоксические клетки – Т-киллеры связываются с антигенами на поверхности клеток и, выделяя белок перфорин, разрушают их. При этом Т-киллер остается жизнеспособным и может разрушать следующую клетку. Действие перфорина подобно МАК системы комплемента. Белок перфорин, полимеризуясь в мембране клетки-мишени, образует поры – каналы, тем самым, вызывая её осмотический лизис. Кроме того, цитотоксический Т-лимфоцит через пору, образованную перфорином в клетке-мишени, вбрасывает гранзимы (ферменты – сериновые протеазы), которые запускают программу апоптоза. Установлено также, что своё цитотоксическое действие Т- лимфоциты могут реализовывать путем экспрессии FasL и с его помощью индуцировать Fas-опосредованный апоптоз мишени.
Тk. Среди субпопуляций Т-лимфоцитов различают эффекторные клетки. В связи с тем, что эти эффекторные клетки способны специфически разрушать клетки-мишени, их называют ЦИТОТОКСИЧЕСКИМИ Т-ЛИМФОЦИТАМИ, или Т-КИЛЛЕРАМИ - убийцами (от англ. to kill - убивать). Т-киллер — одна из основных эффекторных клеток клеточноопосредованного иммунитета, которая наряду с другими клетками способна осуществлять лизис клеток мишеней. Роль Т-киллеров очень велика в реализации трансплантационного иммунитета, развитии аутоиммунных заболеваний, в противоопухолевой защите. Тk-лимфоциты (CD8+ клетки) составляют около 20 – 25% от числа циркулирующих Т-лимфоцитов (абсолютноеколичество - 500 – 1200 в 1 мм3 (мкл)), они несут маркерный антиген CD8. Макромолекула CD8 служит корецептором для антигенов главного комплекса гистосовместимости I класса (ГКГС-1). Активированные антигеном цитотоксические клетки – Т-киллеры связываются с антигенами на поверхности клеток и, выделяя белок перфорин, разрушают их. При этом Т-киллер остается жизнеспособным и может разрушать следующую клетку. Действие перфорина подобно МАК системы комплемента. Белок перфорин, полимеризуясь в мембране клетки-мишени, образует поры – каналы, тем самым, вызывая её осмотический лизис. Кроме того, цитотоксический Т-лимфоцит через пору, образованную перфорином в клетке-мишени, вбрасывает гранзимы (ферменты – сериновые протеазы), которые запускают программу апоптоза. Установлено также, что своё цитотоксическое действие Т- лимфоциты могут реализовывать путем экспрессии FasL и с его помощью индуцировать Fas-опосредованный апоптоз мишени.
«Наивные» Т-лимфоциты – это те лимфоциты, которые не встретились с антигеном и они составляют часть общего пула рециркулирующих Т-клеток.
Т-клетки иммунологической памяти – это долгоживущие лимфоциты, потомки клеток, встречавшихся с антигенами и сохраняющие к ним рецепторы. Т-ЛИМФОЦИТЫ ИММУНОЛОГИЧЕСКОЙ ПАМЯТИ - после стимуляции антигеном способны сохранять информацию о нем до 10—15 лет и передавать ее другим клеткам. Эти клетки защищены от апоптоза. Благодаря наличию в организме Т-клеток памяти обеспечивается ускоренный иммунный ответ по вторичному типу при повторном попадании данного антигена в организм. Этим объясняется форсированная динамика вторичного иммунного ответа. Маркером Т- лимфоцитов памяти является мембранный антиген CD45RO.
Раньше ошибочно выделяли субпопуляцию Т-супрессоров, которые считались ответственными за
подавление иммунного ответа. Однако в настоящее время показано, что самостоятельной субпопуляции Т-супрессоров нет. В угнетении, подавлении иммунного ответа решающее значение играет апоптоз простимулированных лимфоцитов, а также цитокин – трансформирующий фактор роста β.
Около 10 % лимфоцитов не имеют ни Т-, ни В-маркеров, они не относятся ни к Т-, ни к В-лимфоцитам и ранее получили название НУЛЕВЫЕ ЛИМФОЦИТЫ. Эта разнородная популяция лимфоцитов в зависимости от их морфофункциональных особенностей подразделяется на: l ЕСТЕСТВЕННЫЕ (natural) КИЛЛЕРНЫЕ КЛЕТКИ (сокращенно ЕКК= ЕК = NK -клетки) и l КИЛЛЕРНЫЕ КЛЕТКИ (К-клетки). Общей особенностью ЕК- и К-клеток является способность лизировать клетки-мишени без предварительной сенсибилизации, что необходимо Т-лимфоцитам-киллерам. Морфологически это лимфоциты большого размера с зернистой цитоплазмой. Дифференцируются из общей клетки предшественника лимфоцитов (LSC).
 Естественные киллеры не зависят в своем развитии от вилочковой железы. Экспрессируют на своей поверхности рецепторы к интерферону-γ и интерлейкину-2 (ИЛ 2). Функционально они являются цитотоксическими клетками киллерами, но на NK нет антигенраспознающих рецепторов, которые обязательно присутствуют на Т-киллерах. Натуральных киллеров на клетку мишень наводят антитела IgG специфичные к мембранным антигенам клетки-мишени. Первоначально антитела связываются с антигеном на клетке, а затем с помощью Fc рецептора к IgG (FcγRIII) NK присоединяется к этому комплексу АТ - АГ-клетки-мишени. Функция ЕК-клеток в организме заключается в защите от развития опухолей, вирусов и др. Основными их маркерами являются CD16 и CD56. (FcγRIII по CD- номенклатуреэто CD16). Разрушение клетки-мишени ЕК осуществляет с помощью перфорина. Содержание ЕК (CD16+ клеток) у здоровых людей – 8 – 22%.
Естественные киллеры не зависят в своем развитии от вилочковой железы. Экспрессируют на своей поверхности рецепторы к интерферону-γ и интерлейкину-2 (ИЛ 2). Функционально они являются цитотоксическими клетками киллерами, но на NK нет антигенраспознающих рецепторов, которые обязательно присутствуют на Т-киллерах. Натуральных киллеров на клетку мишень наводят антитела IgG специфичные к мембранным антигенам клетки-мишени. Первоначально антитела связываются с антигеном на клетке, а затем с помощью Fc рецептора к IgG (FcγRIII) NK присоединяется к этому комплексу АТ - АГ-клетки-мишени. Функция ЕК-клеток в организме заключается в защите от развития опухолей, вирусов и др. Основными их маркерами являются CD16 и CD56. (FcγRIII по CD- номенклатуреэто CD16). Разрушение клетки-мишени ЕК осуществляет с помощью перфорина. Содержание ЕК (CD16+ клеток) у здоровых людей – 8 – 22%.
К-клетки неоднородная группа клеток, несущая на своей поверхности рецепторы к Fc- фрагменту Ig G и способны к антителозависимой клеточной цитотоксичности. К ним относятся моноциты, нейтрофилы, макрофаги, эозинофилы, конечно НК и некоторые лимфоциты. Антителозависимая
клеточноопосредованная цитотоксичность (АЗКЦ) является своеобразным отражением связи между гуморальным и клеточным звеньями иммунной системы. Антитела выступают в роли «наводчиков» клеток-эффекторов на клетки-мишени, несущие чужеродные антигены.
Все лимфоциты (Т-, В-, ЕК- и К-клетки) обладают способностью к миграции и рециркуляции, что обеспечивает повсеместный контроль за размножением клеток собственного организма, а при проникновении чужеродного антигена —генерализированный иммунный ответ и сохранение иммунологической памяти об антигене.
Date: 2015-08-15; view: 6664; Нарушение авторских прав