Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Кора больших полушарий головного мозга
|
|
Общий план организации коры. Кора больших полушарий является высшим отделом центральной нервной системы, который в процессе филогенетического развития появляется позже всего и формируется в ходе индивидуального (онтогенетического) развития позже других отделов мозга. Кора представляет собой слой серого вещества толщиной 2—3 мм, содержащий в среднем около 14 млрд. (от 10 до 18 млрд.) нервных клеток, нервные волокна и межуточную ткань (нейроглию). На поперечном ее срезе по расположению нейронов и их связей различают 6 горизонтальных слоев. Благодаря многочисленным извилинам и бороздам площадь поверхности коры достигает 0,2 м2. Непосредственно под корой находится белое вещество, состоящее из нервных волокон, которые передают возбуждение в кору и из нее, а также от одних участков коры другим.
Корковые нейроны и их связи. Несмотря на огромное число нейронов в коре, известно очень немного их разновидностей. Основными типами их являются пирамидные и звездчатые нейроны. Которые не различаются по функциональному механизму.
В афферентной функции коры и в процессах переключения возбуждения на соседние нейроны основная роль принадлежит звездчатым нейронам. Они составляют у человека более половины всех клеток коры. Эти клетки имеют короткие ветвящиеся аксоны, не выходящие за пределы серого вещества коры, и короткие ветвящиеся дендриты. Звездчатые нейроны участвуют в процессах восприятия раздражении и объединении деятельности различных пирамидных нейронов.
Пирамидные нейроны осуществляют эфферентную функцию коры и внутрикорковые процессы взаимодействия между удаленными друг от друга нейронами. Они делятся на крупные пирамиды, от которых начинаются проекционные, или эфферентные, пути к подкорковым образованиям, и мелкие пирамиды, образующие ассоциативные пути к другим отделам коры. Наиболее крупные пирамидные клетки — гигантские пирамиды Беца — находятся в передней центральной извилине, в так называемой моторной зоне коры. Характерная особенность крупных пирамид — их вертикальная ориентация в толще коры. От тела клетки вертикально вверх к поверхности коры направлен наиболее толстый (верхушечный) дендрит, через который в клетку поступают различные афферентные влияния от других нейронов, а вертикально вниз отходит эфферентный отросток — аксон.
Многочисленность контактов (например, только на дендритах крупной пирамиды их насчитывают от 2 до 5 тыс.) обеспечивает возможность широкой регуляции деятельности пирамидных клеток со стороны множества других нейронов. Это позволяет координировать ответные реакции коры (в первую очередь ее моторную функцию) с разнообразными воздействиями из внешней среды и внутренней среды организма.
Для коры больших полушарий характерно обилие межнейронных связей. По мере развития мозга человека после его рождения увеличивается число межцентральных взаимосвязей, особенно интенсивно до 18 лет.
Функциональной единицей коры является вертикальная колонка взаимосвязанных нейронов. Вытянутые по вертикали крупные пирамидные клетки с расположенными над ними и под ними нейронами образуют функциональные объединения нейронов. Все нейроны вертикальной колонки отвечают на одно и то же афферентное раздражение (от одного и того же рецептора) одинаковой реакцией и совместно формируют эфферентные ответы пирамидных нейронов.
Распространение возбуждения в поперечном направлении—от одной вертикальной колонки к другой — ограничено процессами торможения. Возникновение активности в вертикальной колонке приводит к возбуждению спинальных мотонейронов и сокращению связанных с ними мышц. Этот путь используется, в частности, при произвольном управлении движениями конечностей.
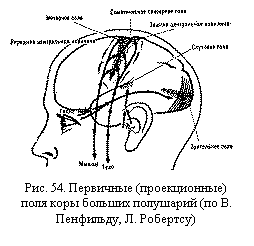 Первичные, вторичные и третичные поля коры. Особенности строения и функционального значения отдельных участков коры позволяют выделить отдельные корковые поля.
Первичные, вторичные и третичные поля коры. Особенности строения и функционального значения отдельных участков коры позволяют выделить отдельные корковые поля.
Различают три основные группы полей в коре: первичные, вторичные и третичные поля.
Первичные поля связаны с органами чувств и органами движения на периферии, они раньше других созревают в онтогенезе, имеют наиболее крупные клетки. Это так называемые ядерные зоны анализаторов, по И. П. Павлову (например, поле болевой, температурной, тактильной и мышечно-суставной чувствительности в задней центральной извилине коры, зрительное поле в затылочной области, слуховое поле в височной области и двигательное поле в передней центральной извилине коры) (рис. 54). Эти поля осуществляют анализ отдельных раздражений, поступающих в кору от соответствующих рецепторов. При разрушении первичных полей возникают так называемая корковая слепота, корковая глухота и т. п. Рядом расположены вторичные поля, или периферические зоны анализаторов, которые связаны с отдельными органами только через первичные поля. Они служат для обобщения и дальнейшей обработки поступающей информации. Отдельные ощущения синтезируются в них в комплексы, обусловливающие процессы восприятия. При поражении вторичных полей сохраняется способность видеть предметы, слышать звуки, но человек их не узнает, не помнит их значения. Первичные и вторичные поля имеются и у человека, и у животных.
Наиболее далеки от непосредственных связей с периферией третичные поля, или зоны перекрытия анализаторов. Эти поля есть только у человека. Они занимают почти половину территории коры и имеют обширные связи с другими отделами коры и с неспецифическими системами мозга. В этих полях преобладают наиболее мелкие и разнообразные клетки. Основным клеточным элементом здесь являются звездчатые нейроны. Третичные поля находятся в задней половине коры — на границах теменных, височных и затылочных ее областей и в передней половине — в передних частях лобных областей. В этих зонах оканчивается наибольшее число нервных волокон, соединяющих левое и правое полушария, поэтому роль их особенно велика в организации согласованной работы обоих полушарий. Третичные поля созревают у человека позже других корковых полей, они осуществляют наиболее сложные функции коры. Здесь происходят процессы высшего анализа и синтеза. В третичных полях на основе синтеза всех афферентных раздражении и с Учетом следов прежних раздражении вырабатываются цели и задачи поведения. Согласно им происходит программирование двигательной деятельности. Развитие третичных полей у человека связывают с функцией речи. Мышление (внутренняя речь) возможно только при совместной деятельности анализаторов, объединение информации от которых происходит в третичных полях.
При врожденном недоразвитии третичных полей человек не в состоянии овладеть речью (произносит лишь бессмысленные звуки) и даже простейшими двигательными навыками (не может одеваться, пользоваться орудиями труда и т. п.).
Воспринимая и оценивая все сигналы из внутренней и внешней среды, кора больших полушарий осуществляет высшую регуляцию всех двигательных и эмоционально-вегетативных реакций.
Функции коры больших полушарии. Кора больших полушарий выполняет наиболее сложные функции организации приспособительного поведения организма во внешней среде. Это прежде всего функция высшего анализа и синтеза всех афферентных раздражении.
 Афферентные сигналы поступают в кору по разным каналам, в разные ядерные зоны анализаторов (первичные поля), а затем синтезируются во вторичных и третичных полях, благодаря деятельности которых создается целостное восприятие внешнего мира. Этот синтез лежит в основе сложных психических процессов восприятия, представления, мышления. Кора больших полушарий представляет собою орган, тесно связанный с возникновением у человека сознания и регуляцией его общественного поведения. Важной стороной деятельности коры больших полушарий является замыкательная функция — образование новых рефлексов и их систем (условные рефлексы, динамические стереотипы—см. главу XV).
Афферентные сигналы поступают в кору по разным каналам, в разные ядерные зоны анализаторов (первичные поля), а затем синтезируются во вторичных и третичных полях, благодаря деятельности которых создается целостное восприятие внешнего мира. Этот синтез лежит в основе сложных психических процессов восприятия, представления, мышления. Кора больших полушарий представляет собою орган, тесно связанный с возникновением у человека сознания и регуляцией его общественного поведения. Важной стороной деятельности коры больших полушарий является замыкательная функция — образование новых рефлексов и их систем (условные рефлексы, динамические стереотипы—см. главу XV).
Благодаря необычайно большой продолжительности сохранения в коре следов прежних раздражении (памяти) в ней накапливается огромный объем информации. Это имеет большое значение для сохранения индивидуального опыта, который используется по мере необходимости.
13. Механизм передачи возбуждения в центральных синапсах, возбуждающие медиаторы, формирование возбуждающего постсинаптического потенциала (ВПСП). Значение хеморегулируемых и потенциалзависимых ионных каналов.
В нервной системе синапсы образуются между отростками разных нейронов, а также между отростками и телами клеток. Количество синапсов на нейроне очень большое и достигает нескольких тысяч. Химические синапсы между нейронами имеют небольшие размеры. Главное отличие нервно-мышечного соединения и межнейронного синапса - тип используемого медиатора. Во всех нервно-мышечных синапсах используется АХ, нейрональные синапсы используют различные медиаторы.
Медиаторами ЦНС являются многие химические вещества, разнородные в структурном отношении (в головном мозге обнаружено около 30 биологически активных веществ). По химическому строению их можно разделить на несколько групп, главными из которых являются моноамины, аминокислоты и полипептиды.
Моноамины – «классические» медиаторы, имеющие в составе 1 атом азота. Это АХ, катехоламины (адреналин, дофамин), серотонин, гистамин. Аминокислоты тоже «классические» медиаторы, глутамат, глицин, ГАМК. Нейропептиды – вещество П, либерины и статины и др. К ним так же относятся эндрфины. Пурины. Газы не являются «класс» медиаторами, посколько не содержатся в синаптических везикулах и не явл стойкими веществами NO, CO, H2S. Для них нет рецепторов на постсин мембранах. Газы диффундируя из нейронов в соседних клетках активируют гуанилатциклазу, фермент, образующий цГМФ, которая запускает процесс фосфолирования белка.
Медиаторы обладают свойством амбивалентности – один и тот же медиатор может активировать как натриевые так и калиевые каналы, т.е. действовать в одних синапсах возбуждающе, а в других вызывать торможнение.
Передача возбуждения в химическом синапсе – сложный процесс в несколько стадий: синтез медиатора, его упаковка в везикулы, прямой аксонный транспорт, секреция в синаптическую щель, взаимодействие с рецепторами постсинаптической мембраны, инактивация избытка медиатора.
Медиаторы обеспечивают одностороннюю передачу возбуждения. Некоторые из них синтезируются в цитоплазме синаптических окончаний, а другие в теле нейрона в аппарате Гольджи (серотонин). В синапсах скелетных мышц – АХ. В синапсах внутренних органов – норадреналин. В синапсах мозга- более 30 медиаторов.
Медиаторы синтезируются постоянно и депонируются в везикулах синаптических окончаний. Синаптические везикулы образуются в теле нервной клетки в ЭПС и цистернах аппарата Гольджи, а затем транспортируются по аксону в нервные окончания.Мелкие синаптические везикулы формируют 2 пула – небольшой, немедленно готовый к экзоцитозу (пул1) и большой (пул2), везикулы которого не участвуют в секреции, но могут транспортироваться в пул 1 и при его истощении пополнять этот пул.
Процессы секреции медиатора с участием синаптических везикул осуществляются в области активной зоны нервного окончания. 5 Этапов секреции:
1. Транспорт везикулы из пула2 в пул1
2. Стыковка везикулы с местом освобождения в активной зоне
3. Подготовка везикулы к экзоцитозу (ожидает входа ионов кальция)
4. Экзоцитоз – слияние мембраны везикулы с плазмотической мембраной (увелич концентр ионов кальция в области везикулы).
5. Рециклизация везикулы посредством эндоцитоза.
Высвобождение медиатора из пузырьков имеет квантовый характер. При возникновении ПД в пресинаптическом нервном окончании деполяризации пресинаптической мембраны высвобождение медиатора резко усиливается. Взаимосвязь между деполяризацией и высвобождением медиатора выполняет Са2+ . Когда ПД достигает пресинаптического полюса кальций входит в цитоплазму пресинаптического окончания и инициирует освобождение медиатора в синаптическую щель.Рецепторы делят на ионотропные и метаботропные.
Взаимодействие медиатора с ионотропным рецептором активирует ионоселективные каналы постсинаптической мемраны. В сост покоя каналы ионотропных рецепторов закрыты. При взаимодействии с медиатором происходит конформационная перестройка и каналы открываются, при этом изменяется проницаемость постсинаптич мембраны для определенных ионов и возникает постсинаптический потенциал.При увеличении проницаемости для натриевых и кальциевых каналов усиливается поступление этих ионов в клетку с последующей деполяризацией химически возбудимых участков мембраны и возникновение ВПСП, величина которого пропорциональна количеству выделившегося медиатора.Из множества ВПСП вследствии их суммации возникает распространяющееся возбуждение в соседнем электровозбудимом участке мембраны (ПД). Такие синапсы называют возбуждающими. Если повышается проницаемость калиевых каналов и каналов для хлора, то происходит снижение возбудимости, передача импульсов затрудняется или прекращается. Такие синапсы являются тормозными.
Метаботропные рецепторы представляют собой сложную систему, состоящую, по крайней мере, из трех белков: 1) собственно рецепторного белка, связывающегося с нейромедиатором; 2) так называемого G-белка, модифицирующего и передающего сигнал с рецепторного белка и 3) белка-эффектора, который является ферментом, катализирующим образование внутриклеточного низкомолекулярного регулятора, так называемого вторичного посредника.
Конечным результатом активации метаботропных рецепторов может быть опосредованное открытие ионных каналов клеточной мембраны, что, в свою очередь, позволяет ионам проникать через них в клетку или, наоборот, выходить через них в межклеточное пространство.
Метаботропные рецепторы не имеют прямой связи с ионными каналами – это самостоятельные белковые структура на поверхности мембраны. Они могут находиться и в синаптической зоне и в любом другом месте на поверхности клетки. Следовательно, на метаботропные рецепторы могут воздействовать, как нейромедиаторы, так и другие лиганды, имеющие внесинаптическое происхождение, например, гормоны, попадающие из кровяного русла в межклеточную жидкость, а затем на метаботропные рецепторы. А нейромедиаторы могут воздействовать на метаботропные рецепторы не только через синапс, но и дистантно, распространяясь диффузно из синаптической щели по межклеточной жидкости.
Инактивирование медиатора необходимо для реполяризации постсинаптической мембраны и восстановления исходного уровня МП. Наиболее важн ым путем инактивирования АХ является ферментативный гидролиз медиатора с помощью ацетилхолинэстеразы. Так же осуществляется удаление медиатора обратным захватом.
Обратный захват из синаптической щели и обратный аксонный транспорт особенно выражен для катехоламинов. Норадреналин и адреналин незначительно разрушаютсяпод действием ингибиторов, поэтому они снова и снова депонируются в цитоплазме пресинаптических окончаний.
14. Виды торможения в центральной нервной системе. Тормозные медиаторы. Механизмы пре- и постсинаптического торможения.
Торможение является одним из фундоментальных свойств ЦНС. И.М. Сеченов обнаружил подавление активности спинного мозга при раздражении структур среднего мозга.
Торможением называется такое синаптическое влияние, которое предотвращает или прекращает возбуждение постсинаптической клетки. Торможение играет важную роль в ЦНС, ограничивая избыточное возбуждение в нейронных сетях.
Выделяют два типа торможения:
1) первичное. Для его возникновения необходимо наличие специальных тормозных нейронов. Торможение возникает первично без предшествующего возбуждения под воздействием тормозного медиатора. Различают два вида первичного торможения:
а) пресинаптическое в аксо-аксональном синапсе;
б) постсинаптическое в аксодендрическом синапсе.
2) вторичное. Не требует специальных тормозных структур, возникает в результате изменения функциональной активности обычных возбудимых структур, всегда связано с процессом возбуждения. Виды вторичного торможения:
а) запредельное, возникающее при большом потоке информации, поступающей в клетку. Поток информации лежит за пределами работоспособности нейрона;
б) пессимальное, возникающее при высокой частоте раздражения;
в) парабиотическое, возникающее при сильно и длительно действующем раздражении;
г) торможение вслед за возбуждением, возникающее вследствие снижения функционального состояния нейронов после возбуждения;
Термины «условное» и «безусловное» торможение предложены И. П. Павловым.
Условное, или внутреннее, торможение — форма торможения условного рефлекса, возникающее при неподкреплении условных раздражителей безусловными. Условное торможение является приобретенным свойством и вырабатывается в процессе онтогенеза. Условное торможение является центральным торможением и ослабевает с возрастом.
Безусловное (внешнее) торможение — торможение условного рефлекса, возникающее под действием безусловных рефлексов (например, ориентировочного рефлекса). И. П. Павлов относил безусловное торможение к врожденным свойствам нервной системы, то есть безусловное торможение является формой центрального торможения.
Пресинаптическое торможение осуществляется тормозными нейронами,, аксоны,которых оказываются на возбуждающих нервных окончаниях персинаптического нейрона образуя аксо-аксональные синапсы. Тормозной медиатор (обычно глицин), выделяющийся в этом синапсе, увеличивает проходимость Cl- и вызывает гиперполяризацию мембраны возбуждающего нервного окончания. Это приводит к снижению возбудимости и увеличению порога генерации ПД пресинаптического нервного окончания.Проведение возбуждения по пресинаптическим путям к постсинаптической мембране блокируется (он до нее не доходит).
Постсинаптическое торможение имеет наибольшее распространение в ЦНС. Оно возникает на постсинаптическом нейроне по механизмам возникновения ТПСП. В этих механизмах участвуют специальные тормозные вставочные нейроны, аксоны которых образуют на соме и дендритах клеток нервных окончаний синапсы(на постсинаптической мембране), выделяя тормозные медиаторы - ГАМК и глицин. Они повышаютпроводимость мембраны для инов Cl- и K+, вызывающих гиперполяризацию постсинаптич мембраны, понижают возбудимость клетки и препятствуют образованию ПД. (Клетки Реншоу или клетки Пуркинье).
Единственными примерами истинно «тормозных «медиаторов являются ГАМК (гамма амино масляная кислота) и глицин.
γ-Аминомасляная кислота (ГАМК, GABA) — аминокислота, важнейший тормозной нейромедиатор центральной нервной системы. Гамма-аминомасляная кислота синтезируется только в нервной системе из глутаминовой кислоты при посредстве глутаматдекарбоксилазы. γ-Аминомасляная кислота выполняет в организме функцию ингибирующего медиатора центральной нервной системы. При выбросе ГАМК в синаптическую щель происходит активация ионных каналов, приводящая к подавлению нервного импульса. Лиганды рецепторов ГАМК рассматриваются как потенциальные средства для лечения различных расстройств психики и центральной нервной системы, к которым относятся болезни Паркинсона и Альцгеймера, расстройства сна (бессонница, нарколепсия), эпилепсия. Известно два типа ГАМК-рецепторов на постсинаптической мембране: ГАМКА (открывает каналы для ионов хлора) и ГАМКБ (открывает в зависимости от типа клетки каналы для. К+ или Са++).
Глицин – аминоускусная кислота, тормозной медиатор в ЦНС. Дополняет активность ГАМК в спинном мозге и стволовых структурах: обеспечивает возвратное торможение мотонейронов, защищая их от перевозбуждения глициновые нейроны – интернейроны моторных ядер. Глициновый интернейрон включается (т.е. Ацх-синапс, образованный коллатералью мотонейрона, запускает в нем ПД) при избыточно сильной активации мотонейрона.
Выделяемый интернейроном глицин вызывает ТПСП на мембране мотонейрона и защищает его от перевозбуждения, а двигательную систему в целом – от судорог
15. Нервно-рефлекторная регуляция физиологических функций. Определение понятия рефлекса, классификация рефлексов. Схема рефлекторной дуги.
Нервная физиологическая регуляция для переработки и передачи информации опосредуется через центральную и периферическую нервную систему. Сигналы передаются с помощью нервных импульсов.
Особенности нервной регуляции:
- имеет точного адресата – сигналы доставляются к строго определенным органам и тканям;
- большая скорость доставки информации – скорость передачи нервного импульса – до 120 м/с;
- кратковременность действия.
Основное положение рефлекторной теории заключается в утверждении, что деятельность организма есть закономерная рефлекторная реакция на стимул. Рефлекс как универсальная форма взаимодействия организма и среды есть реакция организма, возникающая на раздражение рецепторов и осуществляемая с участием центральной нервной системы. И.П. Павлов экспериментально обосновал взгляд на рефлекс как основной акт любой нервной деятельности. Он же разделил все рефлексы по механизму
образования на безусловные и условные.
Безусловные рефлексы – генетически наследуемые, врожденные стереотипные реакции организма на действие внутренних и внешних раздражителей, большинство из них начинает функционировать сразу после рождения. Эти рефлексы характеризуются видовыми особенностями, составляя основу сложных врожденных форм поведения. С их помощью организм совершает различные приспособительные акты в окружающей среде. Более совершенные врожденные безусловные рефлексы называются инстинктами. Без рефл делят на:
- Витальные (жизненно важные) – ориентировочный, рефлекс экономии сил, пищевой, оборонительный.
- Ролевые – родительский, территориальный инстинкты, феномен сопереживания. Реализация их требует взаимодействия с особями того же вида.
- Рефлексы Саморазвития – рефлексы свободы, имитационные, игровые, поисковые ит.д.
Условные рефлексы – индивидуальные приспособительные реакции, приобретенные в течении жизни, формируемые за счет временных связей между сигнальным раздражителем и ответной реакцией организма. С помощью условных рефлексов организм адаптируется во внешней среде. Приспособительный характер условного рефлекса выражается в том, что условный стимул опережающее отражает свойства сочетанного с ним безусловного рефлекса. У человека он может вырабатываться не только на конкретное явление, но и на смысловое содержание слова, обозначающего это явление или раздражитель (вторая сигнальная система).
По биологическому значению выделяют пищевые, оборонительные, питьевые условные рефлексы.
По характеру ответного реагирования: двигательные, секреторные, сердечно-сосудистые.
По степени сложности: простые, комплексные, цепные.
Морфологической основой любого рефлекса является рефлекторная дуга или рефлекторный путь. Рефлекторная дуга (РД) это путь прохождения рефлекторной
реакции, т.е. нервных сигналов.
В естественных условиях рефлекторная реакция происходит при пороговом, надпороговом раздражении входа рефлекторной дуги — рецептора.
Рефлекторная дуга соматического (двигательного) рефлекса состоит из следующих основных звеньев:
1. Рецептор, воспринимающий раздражение
2. Афферентное или восходящее или чувствительное нервное волокно
3. Нервный центр в Ц.Н.С.
4.Эфферентное или нисходящее, двигательное нервное волокно
5. Исполнительный орган “эффектор”
(6.) Обратная связь
Рецептор – это структура, воспринимающая информацию. Рецепторы воспринимают энергию раздражителя и трансформируют ее в энергию нервного импульса. Рецептивные поля разных рефлексов имеют определенную локализацию, рецепторные клетки — соответствующую специализацию для оптимального восприятия адекватных раздражителей (например, фоторецепторы располагаются в сетчатке; волосковые слуховые рецепторы — в спиральном (кортиевом) органе; проприорецепторы — в мышцах, в сухожилиях, в суставных полостях; вкусовые рецепторы на поверхности языка; обонятельные — в слизистой оболочке носовых ходов; болевые, температурные, тактильные рецепторы в коже и т. д.
Классификация рецепторов по виду воспринимаемой информации:
- механорецепторы – воспринимают механическое возбуждение
- терморецепторы – воспринимают температуру
- хеморецепторы – реагируют на химические вещества
- ноцицепторы – болевые рецепторы.
Афферентный путь – дендриты (отростки) чувствительных нейронов. Передает возбуждение от рецепторов в рефлекторный нервный центр.
Рефлекторный нервный центр – совокупность нейронов, расположенных на различных уровнях ЦНС и отвечающих за выполнение сложной рефлекторной функции.
Эфферентный путь представляет собой аксоны нейронов, передающие информацию от рефлекторного нервного центра к рабочему органу.
Эффектор – исполнительный орган, который в ответ на раздражение изменяет свою деятельность. Органами-эффекторами являются мышца или железа.
Обратная связь – это поток импульсов от рецепторов рабочего органа в ЦНС. Он несет информацию об эффективности ответной реакции. За счет обратной связи рефлекторная дуга замыкается в кольцо.
16. Понятие о сенсорных системах: структура и роль. Классификация рецепторов. Рецепторный и генераторный потенциалы. Кодирование сенсорной информации на уровне рецепторов.
Для обеспечения нормальной жизнедеятельности организма необходимы постоянство внутренней среды, связь с непрерывно меняющейся внешней средой и приспособление к ней. Информацию о состоянии внешней и внутренней сред организм получает с помощью сенсорных систем. По назначению:
1. Внешние сенсорные системы – зрительная, слуховая, обонятельная, вкусовая – воспринимают и анализируют изменения параметров окружающей среды.
2. Внутренние (висцеральные) сенсорные системы воспринимают и анализируют изменения параметров внутренних сред организма. Информация из внутренней среды играет важную роль в регуляции функций внутренних органов.
3. Сенсорные сиситемы, оценивающие положение тела - вестибулярная и двигательная – воспринимают и анализируют изменения положения тела в пространстве и частей тела относительно друг друга.
4. Болевая сенсорная система – имеет особое значение для организма- она несет информацию о повреждающих воздействиях.
Анализатор – совокупность центральных и периферических образований, воспринимающих и анализирующих изменения окружающей среды и внутренних сред организма. Любой из анализаторов имеет три отдела:
1. Периферический отдел представлен рецепторами, назначение которых – восприятие и первичный анализ изменений окруж среды и внутренних сред организма. В рецепторах энергия раздражителя трансформируется в нервные импульсы, а так же усиливается сигнал за счет внутренней энергии метаболических процессов.
2. Проводниковый отдел включает цепь афферентных и промежуточных нейронов стволовых и подкорковых структур ЦНС. Проведение возбуждения осуществляют 2 афферентных:
· Специфический проекционный путь (прямые афферентные пути) от рецептора по строго специализированным путям с переключением на различных уровнях ЦНС- спином и продолговатом мозге, в зрительных буграх таламуса и соответствующей проекционной зоне коры большого мозга.
· Неспецифический путь включает РФ, которая на уровне ствола мозга получает от спецефических путей многочисленные коллатерали. За счет коллатералей в процесс возбуждения включается гипоталамус и различные отделы лимбической системы мозга, а так же двигательные центры.
3. Центральный (корковый) отдел состоит из двух частей: центральной - «ядра» спецефических нейронов, перерабатывающих афферентную импульсацию от рецепторов; и периферической - «рассеянных элементов» - нейронов рассредоточенных по коре большого мозга - «сенсорные зоны». В коре входные и выходные элементы образуют колонки – функциональные еденицы коры, организованные в вертикальном направлении. Соседние колонки имеют взаимосвязи, организующие участие множества колонок для осуществления той или иной реакции. Возбуждение одной из колонок приводит к торможению соседних.
К общим принципам организации СС относят:
· Многоуровневость обеспечивает специализацию разных уровней и слоев ЦНС для переработки отдельных видов информации
· Многоканальность проялвяется в наличии параллельных нейрнных путей, обеспечивающих точность анализа воздействующего фактора
· Иерархический принцип – регулирующее нисходящее влияние более высоких уровней на более низкие.
Классификация и механизмы возбуждения рецепторов
Рецепторами называются специальные образования, трансформирующие (преобразующие) энергию внешнего раздражения в специфическую энергию нервного импульса.
Все рецепторы но характеру воспринимаемой среды делятся на эксорецепторы, принимающие раздражения из внешней среды, (рецепторы органов слуха, зрения, обоняния, вкуса, осязания), интерорецепторы, реагирующие на раздражения из внутренних органон, и проприорецепторы, воспринимающие раздражения из двигательного аппарата (мышц, сухожилий, суставных сумок).
По виду воспринимаемых раздражении различают хеморецепторы (рецепторы вкусовой и обонятельной сенсорных систем, хеморецепторы сосудов и внутренних органов); механорецепторы (проприорецепторы двигательной сенсорной системы, барорецепторы сосудов, рецепторы слуховой, вестибулярной, тактильной и болевой сенсорных систем); фоторецепторы (рецепторы зрительной сенсорной системы) и терморецепторы (рецепторы температурной сенсорной системы кожи и внутренних органов).
По характеру связи с раздражителем различают дистантные рецепторы, реагирующие на сигналы от удаленных источников и обусловливающие предупредительные реакции организма (зрительные и слуховые) и контактные, принимающие непосредственные воздействия (тактильные и др.).
По структурным особенностям различают первичные и вторичные рецепторы. Первичные рецепторы — это окончания чувствительных биполярных клеток, тело которых находится вне ЦНС, один отросток подходит к воспринимающей раздражение поверхности, а другой направляется в ЦНС (например, проприорецепторы, терморецепторы, обонятельные клетки). Вторичные рецепторы представлены специализированными рецетпорными клетками, которые расположены между чувствительным нейроном и точкой приложения раздражителя (например, фоторецепторы глаза). В первичных рецепторах энергия внешнего раздражителя непосредственно преобразуется в нервный импульс в одной и той же клетке. В периферическом окончании чувствительных клеток при действии раздражителя возникает повышение проницаемости мембраны и ее деполяризация, возникает местное возбуждение — рецепторный потенциал, который, достигнув пороговой величины, обусловливает появление потенциала действия, распространяемого по нервному волокну к нервным центрам.
Во вторичных рецепторах раздражитель вызывает появление рецепторного потенциала в клетке-рецепторе. Ее возбуждение приводит к выделению медиатора в пресинаптической части контакта клетки-рецептора с волокном чувствительного нейрона. Местное возбуждение этого волокна отражается появлением возбуждающего постсинаптического потенциала или так называемого генераторного потенциала. При достижении порога возбудимости в волокне чувствительного нейрона возникает потенциал действия, несущий информацию в ЦНС. Таким образом, во вторичных рецепторах одна клетка преобразует энергию внешнего раздражителя в рецепторный потенциал, а другая — в генераторный потенциал и потенциал действия. Таким образом, реакция рецепторного нейрона, предназначенного для передачи информации из области восприятия, имеет 5 стадий: 1) преобразование сигнала внешнего раздражения; 2) генерация рецепторного потенциала; 3) распространение рецепторного потенциала по нейрону; 4) возникновение генераторного потенциала; 5) генерация нервного импульса.
Кодирование – преобразование инфы в условную форму (код), удобную для передачи по каналу связи. Характеристики раздражителя кодируются во всех отделах сенсорной системы.
В периферическом отделе кодирование качества раздражителя осуществляется за счет специфичности рецепторов (адекватный раздражитель). Сила раздражителя кодируется изменением частоты импульсов, генерируемых рецепторами. Пространство кодируется величиной площади на которой возбуждаются рецепторы. Время действия раздражителя кодируется тем, что он начинает возбуждаться с началом действия раздражителя, прекращает возбуждаться сразу после выключения раздражителя.
В проводниковом отделе кодирование осуществляется только на «станциях переключения», т.е при передаче сигнала от одного нейрона к другому, где происходит смена кода.
В корковом представительстве происходит частотно-пространственное кодирование. В нейронах коры информация перекодируется в структурные и биохимические изменения (механизмы памяти). В коре мозга осуществляется высший анализ и синтез поступившей информации. Анализ заключается в том, что с помощью ощущений человек различает действующие раздражители по их качеству, определяет силу, время и место, а так же локализацию раздражителя. Синтез реализуется в узнавании известного предмета, явления или в формировании образа впервые встречаемого предмета или явления.
17. Зрительный анализатор. Светопреломляющие среды глаза. Рефракция, ее аномалии и коррекция. Понятие об остроте зрения. Механизмы аккомодации глаза.
С помощью зрительной сенсорной системы воспринимается 80-90 % всей инфы. Благодаря зрению человек может различать освещенность предметов, их цвет, величину, расстояние, на которое они отдалены друг от друга.
Органом зрения является глаз. Орган зрения представлен глазным яблоком и вспомогательными органами – веками, ресницами, конъюктивой и слезным аппаратом, расположенными в глазнице и вокруг нее. Склера – толстая наружная соеденительнотканная оболочка; формирует глазной белок; спереди продолживается в прозрачную мембрану – роговицу. Склера защищает ядро глаза, поддерживает его форму.
Светопремломляющий аппарат глаза построен по принципу последовательно расположенных оптических линз. Роговица является первой и основной преломляющей средой, между ней и радужкой глаза находится передняя камера глаза – полость, заполненная прозрачной жидкостью, выполняющей роль выпукло-вогнутой линзы. В радужной оболочке находится зрачок – отверстие, через которое световые лучи попадают внутрь глазного яблока. Следущая преломляющая среда – хрусталик – двояковыпуклая линза, от кривизны которой зависит фокусировка изображения на сейчастке. За хрусталиком находится основная желеобразная белкосодержащая масса глаза – стекловидное тело.Сохраняет шаровидную форму глаза и удерживает сейчатку в плотном контакте с сосудистой оболочкой и склерой. Сейчатка формирует внутреннюю оболочку глазного яблока.
Периферический отдел сенсорной системы предстален палочковыми и колбочковыми нейросенсорными клетками сейчатки (фоторецепторами). У человека 6-7 млн колбочек и 110-125 млн палочек. Место выхода зрительного нерва из сейчатки – слепое пятно – не содержит фоторецепторов.В области центральной ямки лежит участок наилучшего видения – желтое пятно, содержащее приемущественно колбочки. К периферии сейчатки число колбочек уменьшается, а число палочек возрастает. Периферия сейчатки содержит одни палочки. Палочки воспринимают световые лучи в условиях слабой освещенности (обеспечивают бесцветное зрение). Колбочки функционируют в условиях яркой освещенности и обеспечивают цветное зрение. Фоторецепторы обладают очень высокой чувствительностью (1-2 квантов света достаточно для возбуждения).
В рецепторных клетках сетчатки находятся светочувствительные пигменты – хромопротеиды, которые обесцвечиваются на свету. В палочках – родопсин (сине-зеленый), в колбочках – йодопсин (желт) и др. Под воздействием световой энергии в фоторецепторах происходит сложный молекулярный процесс с участием вторичных посредников, приводящий к распаду зрительных ферментов с последующей их регенерацией при участии витамина А. Этот процесс способствует трансформации энергии в нервные импульсы. В темноте происходит ресинтез пигментов, протекающий с поглощением энергии.
В следствии фотохимических процессов в фоторецепторах сейчатки при действии света возникает рецепторный потенциал, который является следствием гиперполяризации мембраны рецептора. Затем происходит передача сигнала на биполярные клетки и передача нервного импульса по зрительному нерву.
Проводниковый отдел начинающийся в сейчатке (1 нейрон –биполярный, 2 нейрон – ганглиозная клетка) представлен далее зрительными нервами и после частичного перекреста их волокон – зрительными трактами. В каждом зрительном тракте содержатся нервные волокна, идущие от внутренней(носовая) поверхности сейчатки глаза и от наружной половины сейчатки другого глаза. Волокна зрительного тракта направляются к зрительному бугру (собственно таламус). Здесь расположен 3 нейрон от которого нервные волокна уже направляются в КБП.
Центральный отдел (4 нейрон) расположен в затылочной доле КБП. Первичная проекционная область (поле 17), осуществляет более сложную переработку информации. В каждом участке коры сконцентрированы нейроны, которые образуют колонку, проходящую по г
Лубине через все слои вертикально. Разные свойства зрительных объектов обрабатываются в разных частях зрительной коры большого мозга паралленльно. Анализ зрительной инфы не завершается в полях зрителной коры. От поля 17 начинаются пути к другим областям, обеспечивающим не только восприятие зрительного стимула, но и формирование целостного зрительного образа.
Рефракция. Преломляющая сила глаза выражается в диоптриях. Преломляющая сила без явления аккомодации равна 58-60 Д и называется рефракцией глаза. При нормальной рефракции лучи от далеко расположенных предметов после прохождения через светопреломляющую систему глаза собираются на сейчатке в центральной ямке. Моипия (близорукость) – нарушение рефракции, при котором лучи от предмета после прохождения ч\з светпр. Апп. Фокусируются не на сейчатке, а впереди нее. Близкие предметы человек видит без аккомодации, отдаленные – неясными. Нужны очки с рассеивающими двояковогнутыми линзами. Гиперметропия (дальнозоркость) – лучи от далеко расположенных предметов фокусируются за сейчаткой, в силу слабой преломляющей способности глаза. Нужны двояковыпуклые линзы.
Острота зрения определяется наименьшим углом зрения, под которым глаз еще способен видеть раздельно две точки. Чем меньше этот угол, тем выше острота зрения. У большинства людей минимальная острота зрения равна 1 минуте. Определяют с помощью таблиц.
Аккомодация является главным механизмом, обеспечивающим ясное видение разноудаленных предметов. Основной механизм аккомодации заключается в непроизвольном изменении кривизны хрусталика, благодаря чему его преломляющая сила может меняться в пределах 10-14 Д. Хрусталик заключен в капсулу, которая по краям переходит в фиксрующую хрусталик связку, соединенную с волокнами ресничной мышца. При сокращении ресничной мышцы хрусталик становится более выпуклым, преломляющая сила глаза увеличивается, и глаз настраивается на видинее близко расположенных предметов. Когда человек смотри вдаль, ресничная мышца расслабляется, хрусталик уплощается.
18. Строение сетчатки глаза. Фоторецепторы, фотохимические процессы при действии света. Трехкомпонентная теория цветного зрения.
Сетчатка — внутренняя оболочка глаза — имеет сложную структуру. В сетчатке различают две части: заднюю — зрительную, воспринимающую световые раздражения, и переднюю, не содержащую светочувствительных элементов. Снаружи сетчатка выстлана пигментным слоем эпителия, который прилежит к сосудистой оболочке. Зрительная часть сетчатки состоит из десяти слоев клеток: наиболее важными являются три слоя, прилегающие к пигментному эпителию, которые содержат светочувствительные рецепторы — палочки (130 млн.) и колбочки (7 млн.). Это фоторецепторы. Световые лучи от рассматриваемых предметов, проникая через зрачок в глаз, действуют на светочувствительные клетки сетчатки и вызывают в них нервное возбуждение. Колбочки предназначены для дневного зрения. Они малочувствительны к слабому освещению. Ими воспринимаются форма, цвет и детали предметов. Палочки воспринимают световые лучи в условиях сумеречного освещения. Желтое пятно, особенно его центральная ямка, состоящая только из колбочек, является местом наилучшего видения. Такое зрение называют центральным. Остальные части сетчатки принимают участие в боковом, или периферическом, зрении. Центральное зрение обеспечивает возможность рассматривать мелкие детали предметов, а периферическое зрение дает возможность ориентироваться в пространстве.
Каждая колбочка центральной ямки соединена через биполярную клетку с отдельной мультиполярной нервной клеткой, что обусловливает наибольшую остроту зрения в центральной ямке. В других участках сетчатки с каждой мультиполярной нервной клеткой соединено несколько колбочек. В отличие от колбочек большое количество палочек (до 200) соединено с одной биполярной клеткой. Возбуждение палочек и колбочек вызывает появление нервных импульсов в связанных с ними волокнах зрительного нерва.
Колбочки же, в отличие от палочек, бывают трех типов:
1. «Синие» (коротковолновые - S) - 430-470 нм. Их 2% от общего числа колбочек.
2. «Зелёные» (средневолновые - M) – 500-530 нм. Их 32%.
3. «Красные» (длинноволновые - L) – 620-760 нм. Их 64%.
В рецепторных клетках сетчатки находятся светочувствительные пигменты – хромопротеиды, которые обесцвечиваются на свету. В палочках – родопсин (сине-зеленый), в колбочках – йодопсин (желт) и др. Под воздействием световой энергии в фоторецепторах происходит сложный молекулярный процесс с участием вторичных посредников, приводящий к распаду зрительных ферментов с последующей их регенерацией при участии витамина А. Этот процесс способствует трансформации энергии в нервные импульсы. В темноте происходит ресинтез пигментов, протекающий с поглощением энергии. Их восстановление происходит в темноте с участием витамина А, который доставляется к рецепторам по кровеносной системе глаза. Йодопсин восстанавливается быстрее родопсина в 530 раз, поэтому при недостатке витамина А, в первую очередь страдает зрение палочек, или сумеречное зрение.
Слой фоторецепторов лежит на слое пигментных клеток, которые содержат пигмент фуксин. Он поглощает свет и обеспечивает чёткость зрительного восприятия.
В следствии фотохимических процессов в фоторецепторах сейчатки при действии света возникает рецепторный потенциал, который является следствием гиперполяризации мембраны рецептора.
Цветовое зрение – способность зрительной сенсорной системы реагировать на изменения длины световой волны формированием ощущения цвета. Восприятие цвета обусловлено в основном процессами, происходящими в фоторецепторах. Наибольшим признанием пользуется трехкомпонентная теория цветового ощущения Ломоносова – Юнга – Гельмгольца – Лазарева, согласно которой в сейчатке глаза имеется 3 вида фоторецепторов колбочек, раздельно воспринимающих красный, зеленый и сине-фиолетовый цвета. Комбинация возбуждения различных колбочек приводит к ощущению различных цветов и оттенков. Равномерное возбуждение 3 видов колбочек дает ощущение белого цвета. Три типа цветочувствительных колбочек названы модуляторами, а колбочки, которые возбуждаются при изменении яркости света – доминаторами.
Наблюдаются аномалии цветного зрения, которые проявляются в виде частичной или полной цветовой слепоты. Людей, вообще не различающих цвета называют ахроматами. Три вида цветовой слепоты:
· Протанопия – слепота на красный свет
· Дейтеронапия – слепота на зеленый цвет
· Тританопия – слепота на сине-фиолетовый цвет
19. Строение и функции слухового анализатора: звукоулавливающий, звукопроводящий и рецепторный отделы. Анализ высоты и силы звука.
С помощью слуха человек ориентируется в звуковых сигналах окружающей среды, определяет источник звука, и его локализацию в пространстве, физические и информационные характеристики. Слух необходим человеку для речевой коммуникации и умения говорить.
Орган слуха.
1) Наружное ухо – за счет ушной раковины обеспечивает улавливание звуков, концентрацию их в направлении наружного слухового прохода и усиление интенсивности звуков. + защитная функция – охраняют барабанную перепонку от механических и температурных воздействий внешней среды.
2) Среднее ухо – звукопроводящий отдел. Представлено барабанной полостью, где расположены 3 слуховые косточки: молоточек, наковальня и стремечко.
От наружного слухового прохода среднее ухо отделено барабанной перепонкой. Рукоятка молоточка вплетена в барабанную перепонку, другой ее конец сочленен с наковальней, которая сочленена со стремечком. Стремечко примыкает к мембране овального окна. Площадь барабанной перепонки > площади овального окна => усиление давления звуковых волн на мембрану овального окна (примерно в 25 раз).
Защитный механизм: мышца, натягивающая барабанную перепонку; мышца, фиксирующая стремечко. Степень сокращения этих мышц зависит от силы звуковых колебаний. При сильных звуковых колебаниях мышцы ограничивают амплитуду колебаний барабанной перепонки и движение стремечка => предохранение рецепторного аппарата во внутреннем ухе от чрезмерного возбуждения и разрушения. Евстахиева труба – соединяет полость среднего уха с глоткой => в барабанной перепонке поддерживается давление = атмосферному => адекватное восприятие звуков.
3) Внутреннее ухо – представлено улиткой – спирально закрученным костным каналом (2,5 завитка), который разделен основной мембраной и мембраной Рейснера на 3 узких части (лестницы).
Верхний канал – вестибулярная лестница. Начинается от овального окна и соединяется с нижним каналом (барабанной лестницей) через геликотрему (отверстие в верхушке) и заканчивается круглым окном.
Оба канала представляют собой единое целое и заполнены перилимфой. Между верхним и нижним – средний канал (средняя лестница). Изолирован и заполнен эндолимфой. Внутри среднего канала на основной мембране –– собственно звуковоспринимающий аппарат – Кортиев орган (с рецепторными клетками, представляющими периферический отдел слухового анализатора).
Над кортиевым органом – текториальная (покровная) мембрана (из соед.тк.). Один ее край закреплен, др. – свободен. Волоски наружных и внутренних волосковых клеток соприкасаются с текториальной мембраной => изменяется проводимость ионных каналов рецепторных (волосковых) клеток => микрофонный и суммационный рецепторные потенциалы. => образуется и выделяется медиатор – ацетилхолин в синаптическую щель рецепторно-афферентного синапса => возбуждение волокна слухового нерва => ПД в нем => трансформация энергии звуковых волн в нервный импульс.
Периферический отдел представлен рецепторными волосковыми клетками кортиева органа. Фонорецепторы – слуховые рецепторы. Относятся к механорецепторам, являются вторичными и представлены внутренними и наружными волосковыми клетками (примерно 3500 внутренних и 20000 наружных волосковых клеток).Расположены на основной мембране внутри среднего канала внутреннего уха.
Проводниковый отдел представлен периферическим биополярным нейроном (1нейрон), расположенный в спиральном ганглии улитки. Волокна слухового нерва, образованные аксонами нейронов спирального ганглия, заканчиваются на клетках ядер кохлеарного окмплекса продолговатого мозга (2 нейрон). Затем волокна идут в медиальное коленчатое тело метаталамуса, где опять происходит переключение (3 нейрон), откуда возбуждение поступает в кору (4 нейрон).
Центральный отдел - Верхняя часть височной доли большого мозга (верхняя височная извилина, поля 41 и 42 по Бродману). Важное значение для функции слухового анализатора имеют поперечные височные извилины (извилины Гешля).
Слуховая сенсорная система дополняется механизмами обратной связи => регуляция деятельности всех уровней слухового анализатора с участием нисходящих путей. Пути – начинаются от клеток слуховой коры => переключение: в медиальных коленчатых телах метаталамуса => в задних (нижних) буграх четверохолмия => в ядрах кохлеарного комплекса. Входя в состав слухового нерва, центробежные волокна достигают волосковых клеток кортиева органа и настраивают их на восприятие отдельных звуковых сигналов.
Восприятие высоты, силы звука и локализации источника звука. Начинается с попадания звуковых волн в наружное ухо, где они приводят в движение барабанную перепонку => колебания барабанной перепонки через систему слуховых косточек среднего уха передаются на мембрану овального окна => колебания перилимфы вестибулярной (верхней) лестницы => через геликотрему колебания передаются барабанной (нижней) лестнице => доходят до округлого окна => смещение его мембраны по направлению к полости среднего уха. Колебания перелимфы передаются также на эндолимфу перепончатого (среднего) канала => колебательные движения основной мембраны (состоящей из отдельных волокон, натянутых как струны у рояля). Звук действует на волокна мембраны (+ рецепторные клетки кортиева органа, расположенные на них) – колебательные движения. При этом волоски рецепторных клеток контактируют с текториальной мембраной, реснички восковых клеток деформируются. Возникает вначале рецепторный потенциал, а затем ПД (нервный импульс), который далее проводится по слуховому нерву и передается в другие отделы слухового анализатора.
Восприятие звуков различной высоты (частоты).Обусловлено тем, что каждое волокно основной мембраны настроено на звук определенной частоты (низкая частота – длинные волокна основной мембраны ближе к верхушке улитки; высокая частота – короткие волокна основной мембраны ближе к основанию улитки).При действии сложного звука происходят колебания различных волокон мембраны.
Теория места (резонансный механизм) – в состояние колебания вступает вся мембрана. НО: максимальное отклонение – только в определенном месте. Увеличение частоты звуковых колебаний => максимальное отклонение основной мембраны смещается к основанию улитки, где располагаются более короткие волокна (у них возможна более высокая частота колебаний).
Возбуждение волосковых клеток именно этого участка мембраны передается в волокно слухового нерва в виде определенного числа импульсов, частота следования которых ниже частоты звуковых волн (лабильность нервных волокон < 800–1000 Гц).Частота воспринимаемых звуковых волн достигает 20000 Гц. Так осуществляется пространственный тип кодирования высоты и частоты звуковых сигналов. При действии тонов примерно до 800 Гц кроме пространственного кодирования происходит еще и временное(частотное) кодирование, пори котором информация передается также по определенным волокнам слухового нерва в виде импульсов, частота следования которых повторяет частоту звуковых колебаний.
Отдельные нейроны на разных уровнях слуховой сенсорной системы настроены на определенную частоту звука (каждый нейрон имеет свой специфический частотный порог, свою определенную частоту звука, на которую реакция нейрона максимальна). => из всей совокупности звуков каждый нейрон воспринимает только определенные узкие участки частотного диапазона => полноценное слуховое восприятие.
Анализ интенсивности звука. Также осуществляется в слуховой сенсорной системе. При этом сила звука кодируется как частотой импульсов, так и числом возбужденных рецепторов соответствующих нейронов. Наружные и внутренние волосковые рецепторные клетки имеют разные пороги возбуждения. Внутренние клетки возбуждаются при большей силе звука, чем наружные.Кроме того, у внутренних клеток пороги возбуждения также различны.=> от интенсивности звука меняются соотношение возбужденных рецепторных клеток кортиева органа и характер импульсации, поступающей в ЦНС. Нейроны слуховой сенсорной системы имеют различные пороги реакций.При слабом звуковом сигнале в реакцию вовлекаются лишь небольшое число более возбудимых нейронов, а при усилении звука возбуждаются нейроны с меньшей возбудимостью.
20. Роль различных структур центральной нервной системы в организации двигательных функций организма. Рефлексы регуляции длины и напряжения мышц.
Выполнение двигательных актов осуществляется обширным комплексом нейронов, расположенных в различных отделах центральной нервной системы. В регуляции фазной (динамической) и позной (статической) активности мышц принимают участие различные структуры мозга – спинной мозг, ствол мозга, мозжечок, таламус, базальные ганглии и кора больших полушарий. Каждый отдел мозга выполняет определенные задачи, или функции, конкретный характер которых зависит от вида мышечной деятельности.
Каждый из нервных центров, входящих в данный комплекс, может воздействовать как на нижележащие, так и на вышележащие центры. При этом между ними могут возникать циклы взаимных влияний от высших центров к низшим и обратно. Наличие циклических взаимосвязей между различными отделами нервной системы позволяет говорить о замкнутой системе регулирования, или замкнутом цикле регулирования.
В процессе двигательной деятельности одновременно возникает множество замкнутых циклов регулирования (например, между корой и ретикулярной формацией, между корой и мозжечком, между мотонейронами спинного мозга и мышцей и др.). Таким образом, система регуляции движения является многоэтажной (многоуровневой) системой.
К стволу мозга относятся продолговатый мозг, мост и средний мозг. Он так же содержит структуры, принимающие участие в регуляции мышечной активности. Это двигательные ядра черепномозговых нервов, вестибулярные ядра, красное ядро, ретикулярная формация, нейроны покрышки четверохолмия (тектум). Двигательные ядра – скопление альфа-мотонейронов. Эти ядра принимают участие в регуляции сократительной активности поперечно-полосатых мышц лица, глотки, языка, глаз, иннервируют жевательную мускулатуру, а также мышцу, натягивающую барабанную перепонку, иннервируют всю мимическую мускулатуру и др. Осуществляются рефлексы: пищевого поведения (глотание, сосание, жевание), защитные рефлексы – рвота, чиханье, кашель, слезоотделение, смыкание век. Ствол мозга принимает непосредственное участие в регуляции позы тела за счет реализации статических и статокинетических рефлексов. Эти рефлексы предназначены для перераспределения мышечного тонуса, благодаря чему сохраняется удобная для человека поза.
Красное ядро расположено в области среднего мозга. Нейроны этого ядра получают информацию от коры головного мозга, мозжечка, а также от базальных ядер. Красное ядро получает необходимую информацию о положении тела в пространстве, о состоянии мышечной системы и кожных покровов, красное ядро вместе с вестибулярными ядрами участвует в регуляции позы.
Ретикулярная формация ствола мозга – это структура, содержащая нейроны и идущая в ростральном (к коре) направлении от спинного мозга к таламусу и коре больших полушарий. Помимо участия в обработке сенсорной информации (неспецифический канал) ретикулярная формация выполняет и функцию двигательной системы. Нейроны ретикулярной формации продолговатого мозга оказывают такое же влияние на нейроны спинного мозга, как и нейроны красного ядра – при своем возбуждении они активируют альфа-мотонейроны сгибателей и тормозят альфа-мотонейроны разгибателей. Нейроны ретикулярной формации моста, наоборот, действуют подобно нейронам вестибулярных ядер, т. е. повышают активность альфа-мотонейронов разгибателей и тормозят активность альфа-мотонейронов сгибателей.
Основные функции мозжечка – регуляция позы и мышечного тонуса, координация медленных движений и рефлексов поддержания позы и равновесия,коррекция быстрых целенаправленных движений, формируемых двигательной корой больших полушарий.
Соматическая НС регулирует длину и напряжение поперечно полосатых мышц, положение в суставе, определяет позу и передвижение тела в пространстве. На уровне Спиного мозга замыкаются рефлекторные дуги спинальных рефлексов:
Ø Регуляция длины и напряжения мышц
Ø Рефлексы избегания вредных воздействий (отдергивание)
Ø Рефлексы положения (позные)
Ø Сухожильные рефлексы (растяжения)
Ø Ритмические рефлексы, проявляющиеся в правильном чередовании противоположных движений (мышцы антогонисты)
Ø Ритмические рефлексы (потирания, почесывания)
Рефлексы регуляции длины и напряжения мышц
Двигательный аппарат отличается наличием очень специализированных и сложно устроенных рецепторных структур, которые могут быть разделены на два основных типа. Рецепторы одного из них (мышечные веретена) расположены в мышцах, а рецепторы другого - в сухожилиях и суставах. Мышечные веретена расположены между мышечными волокнами и одеты капсулой. Внутри каждого веретена также есть интрафузальные мышечные волокна. Важной особенностью мышечных волокон, расположенных внутри веретена, является наличие у них наряду с двигательными окончаниями рецепторных структур.
Двигательные нервные волокна, иннервирующие интрафузальные мышечные волокна относятся к группе А -гамма. Они являются аксонами особых мотонейронов, соответственно также обозначаемых как гамма - мотонейроны. При усилении их активности сила сокращения этих волокон, напряжение и чувствительность веретен к растяжению растут. С проприорецепторов сигнал идет к чувствительному нейрону, а затем и к эфферентному, расположенному в передних рогах спинного мозга. Он отправляет импульс к экстрафузальным волокнам и мышца сокращается.
Сильное растяжение сухожилия мышцы при ее мощном произвольном соращении приводит к расслаблению этой мышцы. Возбуждаются сухожильные органы Гольджи – окончания чувствительных нейронов и воспринимающие раздражение. Органы Гольджи активизируются при увеличении напряжения в мышце. Афферентные волокна, идущие от них входят в спиной мозг и переключаются на вставочные нейроны, которые образуют тормозные синапсы на альфа-мотонейронах иннервирующих экстрафузальные волокна. Такие рефлексы способствуют быстрому расслаблению мышцы.
21. Вегетативная нервная система: строение, влияние на функции внутренних органов. Строение вегетативной рефлекторной дуги, топография нервных центров и ганглиев симпатического и парасимпатического отделов.
Под ВНС понимают комплекс центральных и периферических образований, обеспечивающих согласованную деятельность внутренних органов с целью приспособления организма к измененяющимся условиям окружающей среды. Под контролем автономной системы находятся органы кровообращения, дыхания пищеварения,выделения, размножения, а также обмен веществ и рост. Фактически эфферентный отдел ВНС осуществляет нервную регуляцию функций всех органов и тканей, кроме скелетных мышц, которыми управляет соматическая нервная система.
Анатомически и функционально вегетативная нервная система подразделяется на симпатическую (центры в тороколюмбальном отделе – грудь и поясница), парасимпатическую (центры распологаются в кранеосокральном отделе – голова и крестец) и метасимпатическую. Симпатические и парасимпатические центры находятся под контролем коры больших полушарий и гипоталамических центров.
В симпатическом и парасимпатическом отделах имеются центральная и периферическая части. Центральную часть образуют тела нейронов, лежащих в спинном и головном мозге. Эти скопления нервных клеток получили название вегетативных ядер. Отходящие от ядер волокна, вегетативные ганглии, лежащие за пределами центральной нервной системы, и нервные сплетения в стенках внутренних органов образуют периферическую часть вегетативной нервной системы.
Симпатические ядра расположены в спинном мозге. Отходящие от него нервные волокна заканчиваются за пределами спинного мозга в симпатических узлах, от которых берут начало нервные волокна. Эти волокна подходят ко всем органам.Распространяющееся из симпатических ганглиев возбуждение широко охватывает сразу несколько различных органов, т.е. является генерализованным.СНС оказывает приемущественно активирующие влияния на функции иннервируемых ею органов: усиливает катаболические реакции, силу и частоту сокращения сердца, повышает АД, увеличивает содержание глюкозы в крови,скорость проведения возбуждения, увеличивает объем легочной вентиляции, расширяет зрачки и др. При этом одновременно снижается тонус органов пищеварительного трата, ослабляются процессы всасывания и ферментативного расщепления в кишечнике. Формирует состояния – агрессия, стресс, болевые реакции.
В СНС передача с преганглионарного на ганглионарный нейроны осуществляется с помощью Н-холино-рецепторов, с помощью АХ, а на эффекторы – через различныевиды адренорецепторов с помощью норадреналина СНС представлена на периферии на значитаельном протяжении постганглионарными волокнами типа С, вследствие чего возбуждение на пути от ганглиев к органам распространяется медленнее, чем в парасимпатической системе. Эффекты действия сим сист более продолжительны чем парасим.
Парасимпатические ядра лежат в среднем и продолговатом мозге и в крестцовой части спинного мозга. Нервные волокна от ядер продолговатого мозга входят в состав блуждающих нервов. От ядер крестцовой части нервные волокна идут к кишечнику, органам выделения. Центральные образования парасим части ВНС лежат в ядрах среднего, продолговатого и спинного мозга. Периферические структуры включают нервные волокна и соответствующие ганглии. Преганглионарные парасим волокна обычно длиннее симпатических. Они идут в составе 3, 6, 9 и 10 пар черепных и 2-4 крестцовых спинномозговых нервов и доходят до околоорганных вегетативных узлов: ресничного, крыловидно-небенного, поднижнечелюстного, ушного, сердечного, легочного, пищеводного, желудочного и др. В них залегают тела вторых эфферентных парасим нейронов, чьи постганглионарные аксоны идут к рабочим органам.
ПНС оказывает успокаивающее, расслабляющее действие на большинство функций организма, снижает возбудимость ЦНС и миокарда, уменьшает интенсивность метаболизма, силу и частоту сердечных сокращений, объем легочной вентиляции, температуру тела, снижает содержание глюкозы в крови. Усиливается моторная, секреторная и всасывательная функции пищеварительного аппарата. Парасим влияния доминируют в формировании сна и психологического субъективного чувства удовлетворения.
Медиатор и в преганглионарных и в постганглионарный синапсах является АХ. Преганглионарные волокна типа В (скорость больше чем у сим).
Метасимпатическая нервная система представлена нервными сплетениями и мелкими ганглиями в стенках пищеварительного тракта, мочевого пузыря, сердца и некоторых других органов. Система микроганглионарных образований обеспечивает этим органам способность к автоматизму.
Деятельность вегетативной нервной системы не зависит от воли человека.
Структура дуги вегетативного рефлекса. Дуга вегетативного рефлекса как и соматическая дуга состоит из трех звеньев – чувствительное, вставочное и эфферентное. Но имеет ряд особенностей.
¨ Возбуждение передается с рецепторов внутренних органов, кровеносных сосудов и др, по волокнам аффернетнф
Date: 2015-07-27; view: 1559; Нарушение авторских прав