Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Нервная ткань
|
|
Нервная ткань (textus nervosus) образует центральную нервную систему (головной и спинной мозг) и периферическую нервную систему - нервы, нервные волокна с их концевыми приборами, нервные узлы (ганглии). Нервная ткань состоит из нервных клеток - нейронов (нейроцитов) с особым строением и функцией, и нейроглии, которая выполняет опорную, трофическую, защитную и разграничительную функции.
Нейроцит (нейрон) (neurocdtus) с отходящими от него отростками является структурно-функциональной единицей нервной системы. Ос- новная функция нейрона - это получение, переработка, хранение, проведение и передача информации, закодированной в виде электрических или химических сигналов (нервных импульсов). В нейроне различают тело, где информация обрабатывается, и отростки, отходящие от тела клетки. Один или несколько древовидно ветвящихся отростков, по которым нервный импульс приносится к телу нейрона, называется дендритом. Единственный отросток, по которому нервный импульс направляется от нервной клетки, - аксон, или нейрит. Нервная клетка динамически поляризована, т. е. способна пропускать нервный импульс только в одном направлении - от дендрита к аксону.
В зависимости от числа отростков различают униполярные, или одноотростчатые, биполярные, или двухотростчатые, и мультиполярные, или многоотростчатые нейроны (рис. 30). К биполярным относятся и ложноуниполярные нейроны (рецепторные нейроны спинномозговых и черепных ганглиев), у которых короткий отросток вскоре Т-образно разветвляет- ся на аксон и дендрит.
Размеры тела нервных клеток колеблются в пределах от 4-5 до 130-140 мкм, а длина отростков может достигать 1 м и более. Форма тел нейро- нов, число дендритов, степень их ветвления широко варьируют в зависимости от локализации нейронов и выполняемой ими функции. Так, например,
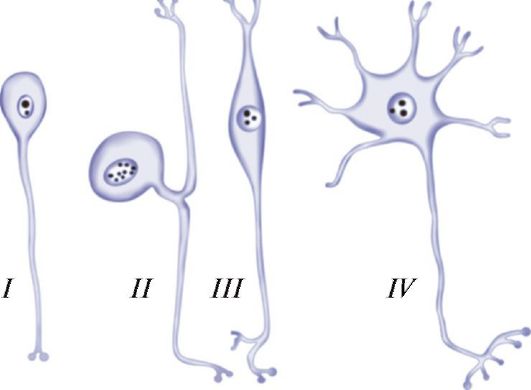
Рис. 30. Типы нейронов: I - униполярный; II - ложноуниполярный; III - биполярный; IV - мультиполярный (по В.Г. Елисееву и др.)
ложноуниполярные клетки спинномозговых узлов имеют округлое тело, ганглионарные мультиполярные нейроны спинного мозга имеют тело неправильной формы, множество слабо ветвящихся дендритов, отходящих в разные стороны, и длинный аксон, от которого отходят коллатерали. От треугольных тел больших пирамидных нейронов коры головного (большого) мозга отходит большое число коротких горизонтальных слабоветвящихся дендритов. Аксон у этих нейронов отходит от основания клетки. В отличие от дендритов, диаметр аксона не меняется на всем протяжении. Грушевидные нейроны коры мозжечка имеют два крупных дендрита, сильно ветвящихся наподобие кроны дерева. Длинный аксон отходит от вершины клетки.
Различают два типа мультиполярных нейронов: длинноаксонные с большим количеством дендритов (клетки типа Гольджи I) и короткоаксонные (клетки типа Гольджи II) с множеством сильно ветвящихся дендритов. К типу I относятся нейроны симпатических и парасимпатических узлов, большие пирамидные нейроны коры головного мозга, грушевидные нейроны коры мозжечка, двигательные нейроны спинного мозга. Эти нейроны передают нервные импульсы на большие расстояния. К типу II относится множество клеток (нейронов) центральной нервной системы, которые передают нервные импульсы многим соседним нейронам.
В сером веществе полушарий большого мозга и мозжечка нейроны располагаются слоями, в других отделах нервной системы образуют скопления (ядра).
Как правило, нейроны являются одноядерными клетками; два ядра имеют некоторые нейроны ганглиев вегетативной нервной системы. Сферическое ядро диаметром около 17 мкм в большинстве нейронов занимает центральное положение (рис. 31). Гетерохроматин располагается по всему ядру. Хорошо выражено базофильное ядрышко, иногда их несколько. В перикарионе находятся многочисленные сферические или удлиненные митохондрии диаметром около 0,1 мкм. Вблизи ядра располагаются десмосомы, связанные между собой элементами комплекса Гольджи. Часто в зоне Гольджи обнаруживаются мультивезикулярные тельца.
Основными особенностями строения нейронов является наличие многочисленных нейрофибрилл и скоплений хроматофильной субстанции (вещество Ниссля), которые представляют собой группы параллельных цистерн зернистой цитоплазматической сети и полирибосомы, богатые РНК. Элементы незернистой эндоплазматической сети в теле клетки малочисленны. Они имеются лишь в аксонах и дендритах в виде трубочек, цистерн и пузырьков. Хроматофильная субстанция и свободные
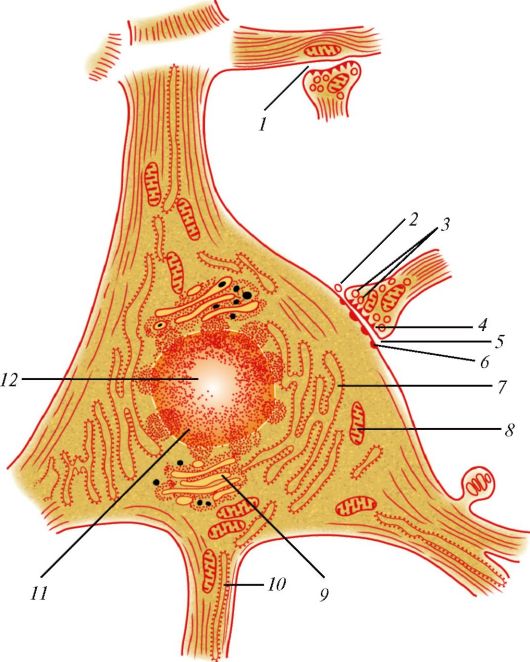
Рис. 31. Схема ультрамикроскопического строения нервной клетки: 1 - аксонодендритический синапс; 2 - аксоносоматический синапс; 3 - преси- наптические пузырьки; 4 - пресинаптическая мембрана; 5 - синаптическая щель; 6 - постсинаптическая мембрана; 7 - эндоплазматическая сеть; 8 - митохондрия; 9 - внутренний сетчатый аппарат (комплекс Гольджи); 10 - нейрофибриллы; 11 - ядро; 12 - ядрышко
рибосомы располагаются по всей цитоплазме клетки и в дендритах, она отсутствует в основании (холмике) аксона и в самом аксоне.
Между элементами эндоплазматической сети расположены многочисленные митохондрии, лизосомы, гранулы липофусцина. Нейрофибриллы, переходящие из тела нейрона в его отростки, образованы микротрубочками диаметром около 20 нм и нейрофиламентами толщиной 7-10 нм.
Нейрофибриллы формируют в перикарионе густую трехмерную сеть, в ячейках которой лежат лизосомы, а также пронизывают цитоплазму отростков. Нейрофибриллы обеспечивают прочность перикариона и от- ростков, осуществляют химическую интеграцию клетки.
Макромолекулы, синтезируемые в перикарионе, направляются в самые отдаленные участки отростков. Посредством постоянного медлен- ного транспорта макромолекул со скоростью 1-3 мм/сут доставляются ферменты, участвующие в синтезе медиаторов в пресинаптической части синапсов, и белки цитоскелета. Быстрый антероградный транспорт со скоростью около 400 мм/сут доставляет пузырьки в синаптические окончания. Кроме того, существует ретроградный транспорт от окончаний аксона к перикариону со скоростью 200-300 мм/сут, с помощью которого более крупные везикулы переносят частицы структур и веще- ства, подлежащие перевариванию в лизосомах. В дендритах также происходит медленный и быстрый транспорт.
В направлении длинной оси дендрита проходит множество нейротрубочек и небольшое количество нейрофиламентов. В цитоплазме дендритов находятся удлиненные митохондрии и небольшое количество цистерн незернистой эндоплазматической сети. Субстанция Ниссля имеется в крупных дендритах, в мелких дендритах элементы незернистой эндоплазматической сети встречаются реже. Конечный отдел дендритов часто колбообразно расширен.
Диаметр аксонов различных клеток (вместе с оболочками) колеблется в широких пределах (от 1 до 20 мкм). Толстые аксоны проводят нервные импульсы быстрее, чем тонкие. Аксоны отходят от конического аксонного холмика, вблизи которого от аксона ответвляются коллатерали. Аксон заканчивается телодендроном - множеством концевых разветв- лений, которые образуют синапсы (контакты). Поверхность цитолеммы аксона (аксолеммы) гладкая. Аксолемма начального сегмента аксона и участка аксона в области узла нервного волокна (перехвата Ранвье) утолщена. В аксоплазме находятся тонкие удлиненные митохондрии, большое количество нейротрубочек и нейрофиламентов, пузырьки и трубочки незернистой эндоплазматической сети, единичные мультивезикулярные тельца. Рибосомы и элементы зернистой эндоплазматической сети отсутствуют в аксоплазме и имеются только в цитоплазме холмика аксона, где расположены пучки микротрубочек, в то время как количество нейрофиламентов здесь невелико.
Итак, нейроны воспринимают, проводят и передают электрические сигналы (нервные импульсы). В основе передачи нервных импульсов лежит изменение мембранного потенциала, вызванного перемещением через
мембрану ионов натрия и калия благодаря функционированию натрийкалиевого насоса.
Нейроны, которые передают возбуждение от точки восприятия раздражения в центральную нервную систему и далее к рабочему органу, связаны между собой с помощью множества межклеточных контактов - синапсов (от греческого syndapsis - связь), передающими нервный импульс от одного нейрона к другому. В синапсах происходит преобразование электрических сигналов в химические и обратно - химических в электрические.
В зависимости от того, какие части нейрона соединены между собой, различают синапсы аксосоматические, когда окончание аксона одного нейрона образует контакты с телом другого; аксодендрические, когда аксоны вступают в контакт с дендритами; а также аксоаксональные, когда контактируют одноименные отростки-аксоны. Такое устройство цепочек нейронов создает возможность для проведения возбуждения от места образования нервных импульсов в спинной или головной мозг и из мозга к рабочим органам (мышцам, железам). При этом передача импульса осуществляется с помощью биологически активных веществ (химическая передача), а сами вещества, осуществляющие передачу, называются нейромедиаторами (от лат. medidator - посредник). Роль медиаторов выполняют норадреналин, ацетилхолин, некоторые моноамины (адреналин, серотонин, дофамин и аминокислоты - глицин, глутаминовая кислота, ГАМК), нейропептиды (энкефалины, нейротензин, ангиотензин II, вазоактивный кишечный пептид, соматостатин, вещество Р и т. д.).
В каждом межнейронном синапсе различают пресинаптическую и постсинаптическую части, разделенные синаптической щелью (рис. 32). Нервный импульс поступает по нервному окончанию в булавовидную пресинаптическую часть нервного окончания, которая ограничена пресинапти- ческой мембраной. Цитоплазма пресинаптической части содержит большое количество округлых мембранных синаптических пузырьков диаметром от 4 до 20 нм, содержащих медиатор. Когда нервный импульс достигает пресинаптической части, открываются кальциевые каналы. Ионы кальция проникает в цитоплазму пресинаптической части, в результате чего его концентрация кратковременно возрастает. При повышении содержания кальция синаптические пузырьки, содержащие нейромедиатор, внедряются в описанные ячейки, сливаются с мембраной, и нейромедиатор выделяется в синаптическую щель, заполненную аморфным веществом умеренной электронной плотности. Чем выше содержание ионов кальция, тем больше синаптических пузырьков выделяют нейромедиаторы. Нейромедиа - тор связывается с рецептором постсинаптической мембраны, что ведет к изменению ее потенциала - возникает постсинаптический потенциал.
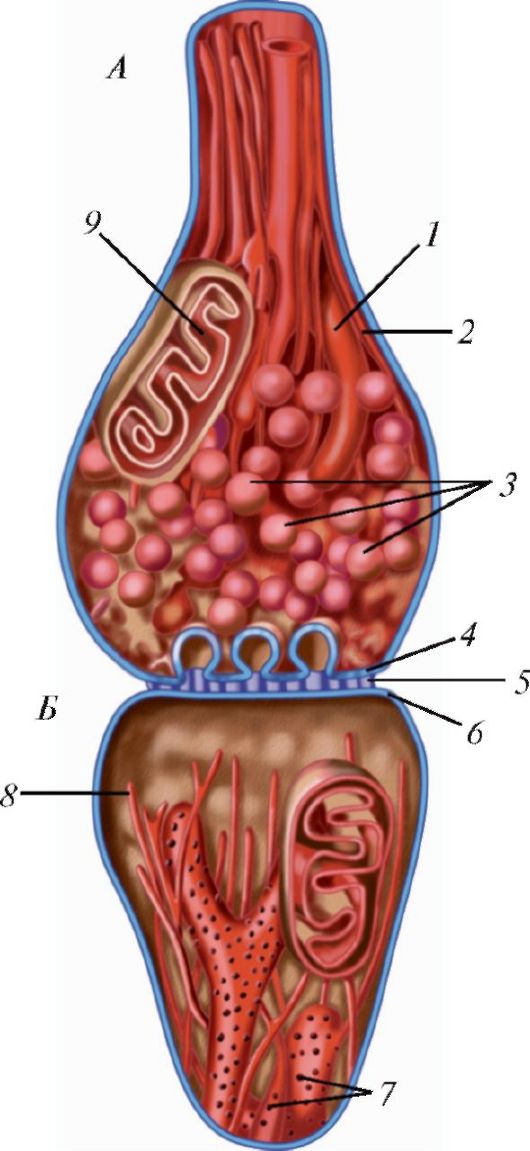
Рис. 32. Схема строения синапса: А - пресинаптическая часть; Б - постсинаптическая часть; 1 - гладкий эндоплазматический ретикулум; 2 - нейротрубочка; 3 - синаптические пузырьки; 4 - пресинаптическая мембрана с гексагональной сетью; 5 - синаптическая щель; 6 - постсинаптическая мембрана; 7 - зернистая эндоплазматическая сеть; 8 - нейрофиламенты; 9 - митохондрия
Таким образом, постсинаптическая мембрана преобразует химический стимул в электрический сигнал. Открываются Na+, К+-каналы, ионы натрия устремляются в постсинаптический полюс, а ионы калия выходят в синаптическую щель, в результате чего происходит деполяризация постсинаптической мембраны. Это ведет к изменению мембранного по- тенциала и возникновению электрического сигнала, величина которого прямо пропорциональна количеству нейромедиатора. Как только прекращается выделение нейромедиатора, пресинаптическое окончание поглощает медиатор из синаптической щели. После этого рецепторы постсинаптической мембраны блокируются антагонистом и возвращаются в исходное состояние.
Нейроглия. Кроме нейронов в нервной системе имеются клетки ней- роглии (neuroglia), выполняющие многообразные функции: опорную,

Рис. 33. Виды нейроглии: I - эпендимоциты; II - протоплаз- матические астроциты; III - волокнистые астроциты; IV - олигоденд- роглиоциты; V - микроглия (по В.Г. Елисееву и др.)
трофическую, защитную, изолирующую, секреторную (рис. 33). Различают две группы нейроглии: глиоциты, или макроглию (эпендимоциты, олигодендроциты и астроциты), и микроглию.
Макроглия. Эпендимоциты (ependymocytus) кубической или призматической формы выстилают изнутри желудочки мозга и спинномозговой канал. Эпендимоциты соединены между собой запирающими зонами и лентовидными десмосомами. От базальной поверхности некоторых эпендимоцитах отходит отросток, который проходит между подлежащими клетками, разветвляется и контактирует с базальным слоем капилляров. Под слоем эпендимоцитов лежит слой недифференцированных глиоцитов.
Астроциты (astrocytus) являются основными глиальными элементами центральной нервной системы. Различают протоплазматические и волокнистые астроциты. Протоплазматические астроциты имеют звездчатую форму, на их телах образуются многочисленные короткие выпячивания, служащие как бы опорой для отростков нейронов, отде- ленных от цитолеммы астроцита щелью шириной около 20 нм. Многочисленные отростки протоплазматических астроцитов заканчиваются на нейронах и на капиллярах. Отростки астроцитов образуют сеть, в которой залегают нейроны. Указанные отростки расширяются на концах, переходя в широкие ножки, которые контактируют между собой. Эти ножки со всех сторон окружают нейроны и кровеносные капилляры, покрывая около 80% их поверхности (вокругсосудистая глиальная пограничная мембрана, membrana limitans glialis perivascularis). He покрыты этой мембраной лишь участки синапсов. Глиальная мембрана, образованная расширенными концами отростков астроцитов, изолирует нейроны, создавая для них специфическое микроокружение. Отростки, достигающие расширенными окончаниями поверхности мозга, соединяясь между собой нексусами, образуют на ней сплошную поверхностную глиальную пограничную мембрану. На этой пограничной мембране лежит базальная мембрана, отграничивающая ее от мягкой мозговой оболочки.
Волокнистые астроциты преобладают в белом веществе центральной нервной системы. Это многоотростчатые (20-40 отростков) клетки, тела которых имеют размеры около 10 мкм. Отростки располагаются между нервными волокнами, некоторые отростки достигают кровеносных капилляров.
Олигодендроциты (oligodendrocytus) - это мелкие клетки овоидной формы (6-8 мкм) с крупным, богатым хроматином ядром, окруженным тонким ободком цитоплазмы, в которой находятся умеренно развитые
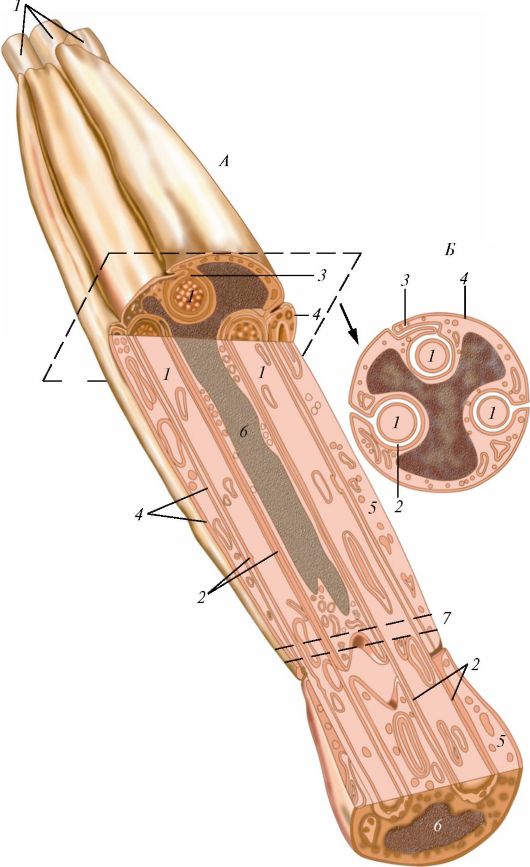
Рис. 34. Схема строения безмиелинового (безмякотного) нервного волокна: А - продольный срез; Б - поперечный срез; 1 - осевые цилиндры; 2 - аксолемма; 3 - мезаксон; 4 - клеточная оболочка нейролеммоцита (шванновской клетки); 5 - цитоплазма нейролеммоцита; 6 - ядро нейролеммоцита; 7 - контакт двух нейролеммоцитов (по В.Г. Елисееву и др.)
органеллы. Олигодендроциты располагаются вблизи нейронов и их отростков. От тел олигодендроцитов отходит небольшое число коротких конусовидных и широких плоских трапециевидных миелинобразующих отростков. Эти отростки формируют миелиновый слой нервных волокон в центральной нервной системе. Миелинобразующие отростки каким-то образом спирально накручиваются на аксоны.
Микроглия (microglia), составляющая около 5% клеток глии в белом веществе мозга и около 18% в сером, состоит из мелких удлиненных клеток угловатой или неправильной формы, рассеянных в белом и сером веществе центральной нервной системы (клетки Ортега). От тела клетки отходят многочисленные отростки, напоминающие кустики. Основания некоторых клеток микроглии как бы распластаны на капилляре.
Нервные волокна (neurofibrae) представляют собой отростки нервных клеток, окруженные оболочками, образованными олигодендроцитами (нейролеммоциты, или шванновские клетки). Различают миелиновые и безмиелиновые волокна. У безмиелиновых волокон, встречающихся преимущественно в нервах вегетативной нервной системы, отростки нейронов (осевые цилиндры) прогибают цитоплазматическую мембрану олигодендроцита (нейролеммоцита), которая окутывает (окружает) эти отростки (рис. 34). В одну шванновскую клетку может быть погружено несколько осевых цилиндров.
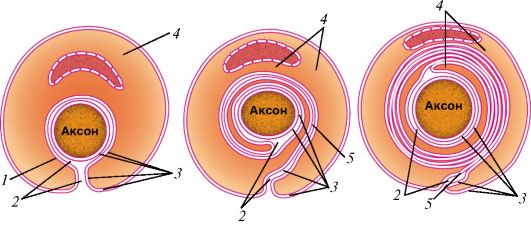
Рис. 35. Формирование миелинового (мякотного) нервного волокна:
1 - контакт аксолеммы и цитолеммы нейролеммоцита (шванновской клетки);
2 - межклеточная щель; 3 - аксолемма и цитолемма нейролеммоцита; 4 - цито-
плазма нейролеммоцита; 5 - мезаксон (по В.Г. Елисееву и др.)
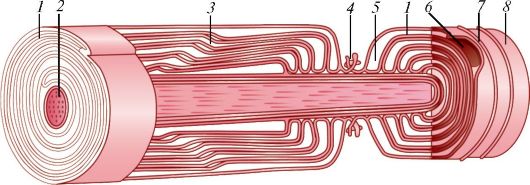
Рис. 36. Строение миелинового (мякотного) нервного волокна: 1 - мезаксон; 2 - осевой цилиндр; 3 - насечки нейролеммы; 4 - узел нервного волокна; 5 - цитоплазма нейролеммоцита; 6 - ядро нейролеммоцита; 7 - нейро- лемма; 8 - эндоневрий (по В.Г. Елисееву и др.)
Миелиновые нервные волокна толще безмиелиновых, образуются благодаря тому, что нейролеммоцит (шванновская клетка) спирально накручивается на осевой цилиндр (аксон) нервной клетки (рис. 35). Миелин - это многократно закрученный двойной слой цитоплазматической мембраны нейролеммоцита. Толстая и плотная миелиновая оболочка, богатая липидами, изолирует нервное волокно и предотвращает утечку нервного импульса из осевого цилиндра. Каждый нейролеммоцит окутывает только часть осевого цилиндра длиной около 1 мм, образуя межузловой сегмент нервного волокна. На границе между двумя соседними нейролеммоцитами образуется сужение нервного волокна - узловой перехват нервного волокна (перехват Ранвье) шириной около 0,5 мкм, где миелиновая оболочка отсутствует (рис. 36). Здесь осевой цилиндр контактирует с цитолеммой нейролеммоцитов.
В зависимости от функции выделяют три основных типа нейронов: афферентные, ассоциативные и эфферентные. Чувствительные, рецепторные, или афферентные, нейроны (от лат. afferdns - приносящий) проводят нервные импульсы от органов и тканей в мозг. Тела таких нейронов у человека лежат вне центральной нервной системы. Как правило, это би- полярные (ложноуниполярные) нейроны. Один из отростков, отходящих от тела чувствительной нервной клетки, следует на периферию и заканчивается тем или иным чувствительном окончанием (рецептором), который способен трансформировать энергию внешнего раздражителя в нервный импульс. Второй отросток направляется в головной или спинной мозг. В зависимости от локализации различают: 1) экстерорецеп- торы, воспринимающие раздражение внешней среды. Они расположены
в наружных покровах тела, в коже и слизистых оболочках, в органах чувств; 2) интерорецепторы, получающие раздражение главным образом при изменении химического состава внутренней среды (хеморецепто- ры) и давления в тканях и органах (барорецепторы, механорецепторы); 3) проприорецепторы, воспринимающие раздражение в мышцах, сухожилиях, связках, фасциях, костях, суставных капсулах и т. д.
В зависимости от характера восприятия раздражения различают терморецепторы, механорецепторы и ноцирецепторы. Терморецепторы воспринимают изменение температуры, механорецепторы улавливают различные виды механических воздействий (прикосновение к коже, ее сдавление), ноцирецепторы воспринимают болевые раздражения.
Среди нервных окончаний различают свободные нервные окончания, не имеющие какой-либо оболочки, и несвободные (инкапсулированные), которые покрыты тонкой соединительнотканной пластинкой или слоем клеток нейроглии (рис. 37).
Свободные (неинкапсулированные) нервные окончания (terminatio nervi libera) имеются в коже. Подходя к эпидермису, нервное волокно теряет миелин, проникает через базальную мембрану в эпителиальный слой кожи. Нервные волокна разветвляются между эпителиоцитами вплоть до зернистого слоя, их веточки диаметром менее 0,2 мкм колбообразно расширяются на концах. Аналогичные концевые нервные окончания имеются в эпителии слизистой оболочки и роговицы глаза. Концевые свободные нервные окончания воспринимают боль, тепло и холод.
В эпидермисе свободные нервные окончания подходят также к измененным эпителиоцитам (клеткам Меркеля), которые имеют пальцевидные выросты. Эти окончания являются механорецепторами, воспри- нимающими давление.
Рис. 37. Схема строения рецепторов: А: 1 - свободное нервное окончание; 2 - граница между дермой и эпидермисом; Б: 1 - измененная клетка эпидермиса (клетка Меркеля); 2 - базальная мембрана; 3 - конечный диск афферентного волокна; 4 - миелин; 5 - нейролеммоцит; В: 1 - субкапсулярное пространство; 2 - капсула; 3 - миелин; 4 - нейролеммоцит; 5 - наружная колба; 6 - базальная мембрана; 7 - внутренняя колба; 8 - терминальный отросток афферентного волокна; Г: 1 - уплощенные нейролеммоциты; 2 - капсула; 3 - базальная мембрана; 4 - спиральные терминали афферентного волокна; 5 - нейролеммоциты; 6 - миелин; Д: 1 - пучки коллагеновых волокон в ядре тельца; 2 - терминальные ветви афферентного волокна; 3 - капсула; Е: 1 - терминальные ветви афферентного волокна; 2 - капсула (по А. Хэму и Д. Кормаку)
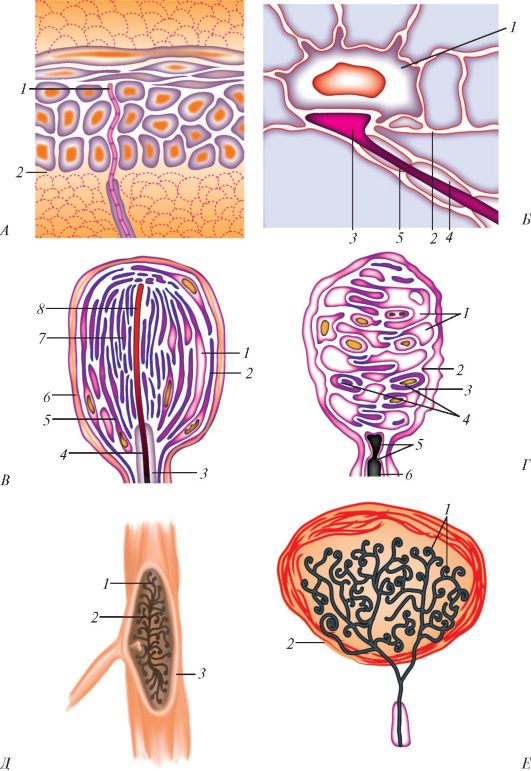
Барорецепторы представляют собой ветвящиеся свободные нервные окончания, залегающие в адвентициальном слое крупных артерий грудной полости и шеи. Наиболее важные из них - рецепторы, залегающие в стенках дуги аорты и сонного синуса. При растяжении стенки артерии под влиянием изменяющегося артериального давления эти окончания возбуждаются. Участие барорецепторов в регуляции артериального давления осуществляется по принципу обратной связи.
Несвободные (инкапсулированные) нервные окончания - это осязательные тельца (Мейсснера), пластинчатые (Фатера-Пачини), луковицеобразные (Гольджи-Маццони). Все эти нервные окончания являются механорецепторами.
Пластинчатые тельца Фатера-Пачини - самые крупные из всех инкапсулированных нервных окончаний. Они имеют овальную форму, длину 3-4 мм и толщину 2 мм, располагаются в соединительной ткани внутренних органов и в подкожной основе (рис. 38). Тельце снаружи покрыто соединительнотканной капсулой, имеющей пластинчатое строение. Под соединительнотканной оболочкой лежит наружная луковица, состоящая из 10-60 концентрических пластинок, образованных уплощенными периневральными эпителиоидными клетками. Войдя в тельце, нервное волокно теряет миелиновую оболочку, формирует внутреннюю луковицу и заканчивается колбообразным вздутием.
Осязательные тельца (Мейсснера) длиной 50-160 мкм, шириной около 60 мкм, овальные или цилиндрические, особенно многочисленны в сосочковом слое кожи пальцев рук, а также в коже стопы. Тельце образовано удлиненными, уплощенными шванновскими клетками, лежащими одна на другой. Нервное волокно, входя в тельце, теряет миелин и оканчивается колбообразным расширением. Периневрий переходит в окружающую тельце капсулу, образованную несколькими слоями эпителиоидных периневральных клеток. Сдавление этих клеток передается осязательному тельцу, вызывая деформацию его клеток, что приводит к возникновению импульса в нервных волокнах. Таким образом, мейсснеровские тельца являются механорецепторами, воспринимающими прикосновение, сдавление кожи.
Концевые колбы (Краузе) расположены в коже, конъюнктиве глаз, слизистой оболочке ротовой полости. Сферические тельца окружены толстой соединительнотканной капсулой, богатой коллагеновыми волокнами и фибробластами. Войдя в капсулу, нервное волокно теряет миелиновую оболочку и разветвляется в центре колбы, образуя множество веточек. Колбы Краузе воспринимают холод, возможно, они являются и механорецепторами.
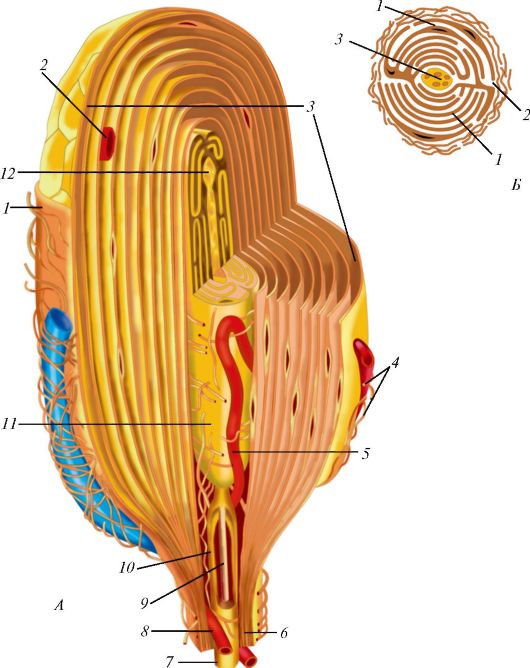
Рис. 38. Схема пластинчатого тельца Фатера - Пачини: А - продольный срез: 1 - соединительнотканная капсула; 2, 5, 8 - гемокапилляры; 3 - пластинки наружной луковицы; 4 - коллагеновые микрофибриллы; 6 - периневрий; 7 - миелиновое нервное волокно; 9 - аксон; 10 - миелин; 11 - внутренняя луковица; 12 - окончание нервного волокна; Б - поперечный срез: 1 - интердигитирующие полулуковицы внутренней луковицы; 2 - радиальная щель; 3 - нервное волокно (по Р. Крстичу, с изменениями)
Проприорецепторы воспринимают тягу сокращения мышц, натяжения сухожилий и суставных капсул, возникающих при выполнении определенного движения или удержания частей тела в определенном положении. Это нервно-мышечные и нервно-сухожильные веретена, которые находятся в брюшке мышц или в их сухожилиях. Сухожильные органы (Гольджи) расположены внутри сухожилий, поблизости от мышц.
Нервно-мышечное веретено крупное (длиной 3-5 мм и толщиной до 0,5 мм), окружено плотной соединительнотканной капсулой, состоящей из внутренней и наружной пластинок, в которую заключены от 2 до 12 мышечных, а также нервных волокон. Продольно расположенные веретена растягиваются вместе с мышцей и несут информацию о длине и степени растяжения мышцы.
Вставочный (ассоциативный), или кондукторный, нейрон осуществляет передачу нервного импульса с чувствительного (центростремительного) нейрона на двигательный или секреторный (центробежный).
Тела эфферентных (эффекторных, двигательных или секреторных) нейронов (от лат. efferens - выносящий) находятся в центральной нервной системе или в симпатических и парасимпатических нервных узлах. Аксоны эфферентных нейронов несут нервные импульсы к рабочим органам (мышцам или железам), а также ко всем органам и тканям для иннервации (регуляции) обмена веществ. Выделяют также двигательные и секреторные
окончания аксонов выносящих (эфферентных) нейронов. Аксоны двигательных нейронов разветвляются и каждый из них ин- нервирует большое количество мышечных волокон. Двигательные (моторные) нервные окончания располагаются на мышечных волокнах. Окончание одного двигательного нервного окончания и иннервируемое им поперечнополосатое мышечное волокно образуют двигательную единицу - мион.
Каждое мышечное волокно иннервируется веточкой аксона двигательного нейрона, который, оканчиваясь на волокне,
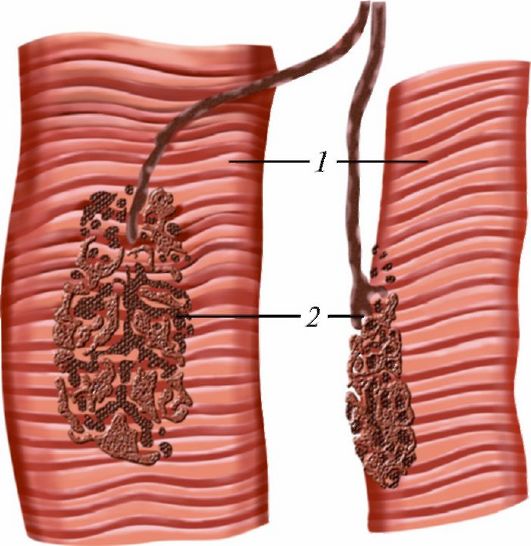
Рис. 39. Нервно-мышечная пластинка: 1 - мышечные волокна; 2 - моторные бляшки
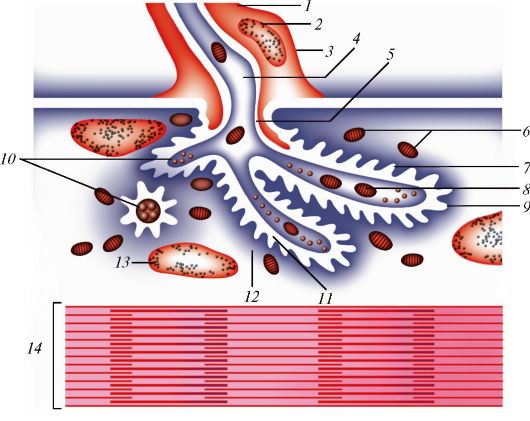
Рис. 40. Строение аксомышечного синапса: 1 - нейролеммоцит; 2 - ядро нейролеммоцита; 3 - нейролемма (цитоплазматическая мембрана нейролеммоцита); 4 - аксоплазма; 5, 11 - аксолемма (пресинаптическая мембрана); 6 - митохондрии мышечного волокна; 7 - синаптическая щель; 8 - митохондрия нервного окончания; 9 - сарколемма (постсинаптическая мембрана); 10 - синаптические пузырьки; 12 - сарколемма; 13 - ядро мышечного волокна; 14 - миофибриллы (по В.Г. Елисееву и др.)
образует нейромышечное окончание, или двигательную концевую пластинку (рис. 39), у которой различают пресинаптическую и постсинаптическую мембраны, разделенные синаптической щелью (рис. 40).
Нервный импульс, достигающий нервно-мышечного окончания, приводит к выходу в синаптическую щель ацетилхолина, который связывается со специфическими рецепторами мембраны мышечного волокна.
Нервные окончания неисчерченной (гладкой) мышечной ткани располагаются между гладкомышечными клетками, где образуют расширения - синаптические пузырьки, плотно прилежащие к базальной мембране миоцитов.
Date: 2015-06-11; view: 2832; Нарушение авторских прав