Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Механизмы поступления воды в клетку
|
|
Клетка как основная структурная и функциональная единица растительного организма, является и основной структурой поглощения воды любых тканей и органов. Поэтому здесь уместно рассмотреть поступление воды в клетку.
Чтобы понять основные механизмы поступления воды в клетку, надо уяснить некоторые физические явления, которые лежать в основе этого физиологического процесса.
Первоначально возникло представление о поступлении воды в клетку диффузией. Диффузия (от лат слова diffusion, что означает расширение, растекание) – это процесс равномерного распределения молекул растворенного вещества и растворителя. Она всегда направлена по градиенту концентрации, от большей концентрации к меньшей, от системы, обладающей большей свободной энергией, к системе с меньшей свободной энергией, независимо это молекулы растворителя или растворенного вещества. Свободная энергия это та, которая может совершать работу. Если её отнести к 1 молю вещества, то она называется химическим потенциалом. Чем выше концентрация вещества, тем большую работу осуществляют его молекулы, тем выше его химический потенциал. Наибольший химический потенциал у чистой воды. Если к воде добавить растворенное в ней вещество, то её химический потенциал уменьшается. Поэтому для воды вводится понятие водный потенциал. Водный потенциал наибольший в химически чистой воде и от падает с растворением в ней какого-либо вещества.
Поэтому водный потенциал чистой воды равен 0 (Ψ ч. воды), а в растворе имеет отрицательную величину. Но диффузия идет очень медленно и может осуществляться в ограниченном пространстве, например, внутри цитоплазмы, в клеточном соке, в свободном пространстве клеточных стенок.
Если на пути между растворителем и раствором поставить мембрану, то она замедлить диффузию, особенно, если она будет пропускать растворитель, и не пропускать растворенное вещество. Такое явление проникновения молекул растворителя через полупроницаемую мембрану, вызванное разностью концентраций, называется осмосом.
Простейшая модель осмометра впервые 1826 году была показана французским ботаником Г. Дютроше. К кончику стеклянной трубки он прикрепил пергаментный мешочек, залил в него раствор соли и поместил в стакан с водой. В мешочек поступала вода и поднималась в стеклянной трубке.
Немецкий ботаник В. Пфеффер в 1877 г создал более совершенную модель осмометра, отражающего модель клетки. Роль клеточной стенки играл пористый фарфоровый сосуд. Налив в фарфоровый сосуд раствор медного купороса и поместив этот сосут в сосуд с фарроционидом меди Сu2[Fe(CN)6. Соединившись эти два раствора в порах фарфрового сосуда, образовали полупроницаемую мембрану, проницаемую для воды и не проницаемую для растворенных в ней веществ.
Затем фарфоровый сосуд заполнили раствором сахарозы, играющего роль клеточного сока, и поместили в сосуд с чистой водой. Вода стала поступать в сосуд, что было видно по поступлению воды в трубку. То же самое наблюдается и в клетке: если поместить её в воду, то вода поступает в вакуоль.
Таким образом было показано, что Более концентрированный раствор – это клеточный сок, менее концентрированный (наружный) находится в свободном пространстве клеточной стенки, а роль полупроницаемой мембраны выполняют сообщаплазмалемма, тонопласт и расположенная между ними цитоплазма.
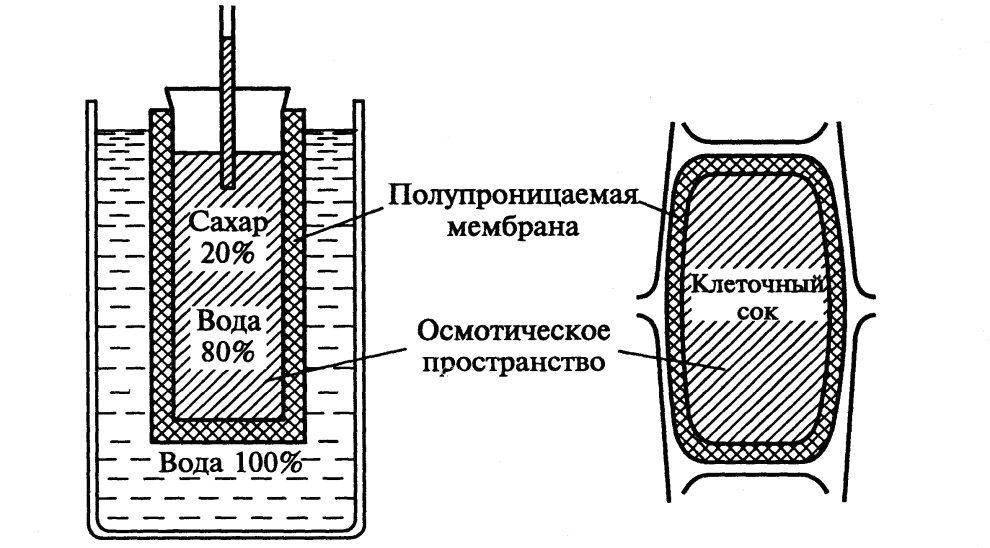
Рис2. Осмометр Пфеффера и растительная клетка (из Э. Либерта. 1976)
.
Пфеффер установил, что в осмометре вода поступающая в клетку поднимается по трубке до тех пор, пока давление водяного столба в ней не станет равной силе, с которой молекулы воды поступают в осмометр. То давление, которое необходимо приложить к раствору, чтобы предотвратить поступление воды через полупроницаемую мембрану, назвали осмотическим давлением. Пфеффер установил, что его величина зависит от концентрации раствора
Датский химик Я. Вант- Гофф, обобщая результаты Пфеффера показал, что осмотическое давление подчиняется законам для идеальных газов и прежде закону Бойля-Мариотта, т.е. зависит от молярной концентрации, т.е. числа растворенных частиц. Любые растворы веществ не электролитов изоосмотические. Согласно правила Авогадро молекулярный раствор имеет осмотическое давление 22,4 атм. или 2,23 МПа (мегапаскаля). Осмотическое давление подчиняется и закону Гуй-Люсака, т.е. возрастает от повышения температуры. Поэтому раствор, находящийся даже в стеклянном сосуде обладает осмотическим давлением, который может проявиться только при конnакте с другим раствором, через полупроницаемую мембрану, а поэтому любой раствор обладает осмотическим потенциалом, так же как и клеточный сок вакуоли. Поэтому осмотическое давление, или осмотический потенциал (Ψ осм) не электролитов (сахароза) рассчитывается по формуле Клапейрона:
Ψосм.= RTc
где К – газовая постоянная, равная 0,082; Т – абсолютная температура; с – концентрация раствора в молях. Для электролитов, который в водном растворе подвергаются электролитической диссоциации на ионы, приводит к образованию в растворе большего числа частиц и этим увеличивает осмотическое давление (NaClNa+ + Cl-). Поэтому в уравнение водится поправка – изотонический коэффициент i, и оно приобретает вид:
Ψосм.= RTci
На величину осмотического давления влияет лишь концентрация растворенных в воде веществ. Эти вещества называются осмотически активными (осмолитиками) К ним относятся органические кислоты, аминокислоты, сахара, соли. Растворы с одинаковым осмотическим давлением называют изотоническими (изоосмотическими); между ними осмос не наблюдается. Раствор, имеющий бόольшее осмотическое давление, называют – гипертоническим, меньшее – гипотоническим. И после работ Пфеффера поступление воды в клетку объясняли только разностью осмотического давления клеточного сока и наружного раствора: если клетка находится в гипотоническом растворе или воде, вода входит клетку, (эндосмос), в гипертоническом – вода выходит из клетки (экзоосмос). Т.Е от величины осмотического давления клеточного сока будет зависеть и поглощение воды из почвы. Осмотическое давление, или осмотический потенциал, у разных растений различен. Это зависит от их экологии и условий произрастания. У водных растений он составляет около 1 атм. (0,1 МПа), у морских водорослей 25-30 атм. (2,5- 3,0 МПа), у мезофитов –3-7 атм.(0,3-0,7 МПа), у ксерофитов 35-60 атм. (3,5 – 6,0 МПа), а у галофитов до 100-200 атм. (10-20 МПа).
В 1918 году а, Уршпрунг и Г.Блюм доказали, что осмотическое давление не является единственным условием поступления волы в клетку. Поступающая вода в клетку увеличивает объём вакуоли, которая давит на цитоплазму в связи с возникающим гидростатическим давлением воды на цитоплазму, которая давит на клеточную стенку и растягивает её. В с вою очередь, растягивающаяся клеточная стенка оказывает противодавление с такой же силой на цитоплазму, которое получило название тургорного давлении (потенциал давления Ψдав.), а напряженное состояние клеточной стенки тургором. Но клеточная стенка не может растягиваться до бесконечности и наступает такое явления, когда вода в клетку уже не поступает, так как оболочка уже не растягивается. Тургорное давление нивелировало всю силу осмотического потенциала. Такое состояние выражается формулой – Ψ осм. = Ψ дал. Следовательно, поступление воды связано не только с осмотическим давлением, но и тургорным давлением. Чтобы вола поступала в клетку, между ними должна быть разница. Разница между осмотическим потенциалом и потенциалом давления и обеспечивает поступление воды в клетку, она получила название сосущая сила.
В1960 году был введен термин «водный потенциал клетки» (Ψ Кл), заменивший понятие сосущая сила .– это разность между свободной энергией воды внутри и вне клетки при той же температуре и атмосферном давлеии. Воднй потенциал клетки – это мера энергии, с которой вода устремляется в клетку. Активность – это производное концентраций. Присутствие растворенных веществ в клетке уменьшает концентрацию воды, снижая её активность, т.е. увеличение снижает её водный потенциал становится более отрицательным (–Ψвод). У слабого раствора Ψвод. = – 0.1, сильно концентрированного –0,9.Таким образом, водный потенциал клетки показывает, насколько энергия воды в клетке меньше энергии чистой воды, и характеризует способность воды диффундировать, испаряться или поглощаться. Так как в клетке при поступлении воды создается гидростатическое давление, растягивающее клеточную стенку, и образуемого её противодавление – тургорное давление потенциал давления), имеющего положительную величину, то водный потенциал выражается формулой:
– Ψ вод. = – Ψ осм. – (+Ψ дав.)
Сосущая сила самая переменная величина и зависит от степени насыщенности клетки водой.. Если клетка насыщена водой и в ней не нуждается, то сосущая сила равна 0, так как – Ψ осм. = +Ψ дав. И такое состояние клетки называется полным насыщением водой. Но в природе такое состояние весьма и весьма редко, так как в процессе испарения (транспирации) Ψдав.<Ψосм. Обычное состояние заключается том, что клетка несколько недонасыщенна из-за того, что идет потеря воды при транспирации, поэтому действуют условия ости между осмотическим потенциалом и потенциалом давления.
| Рис3 Схема изменения осмотических величин в клетке при переходе от завя-дания к насыщению водой (слева на пра-во): Т–тургорное давление; О –отичес- кое давление клеточного сока; S– сосу- щая сила (по Н.А. Максимову) |
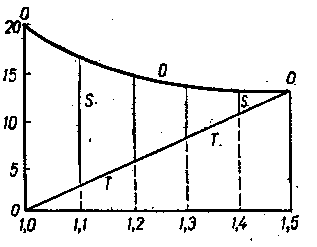 В данном случае растительная клетка выступает как саморегулирующая система, обеспечивающая непрерывное поглощение воды, поддерживая равновесие. Причем поглощение воды тем сильнее, чем больше она в ней нуждается. В состоянии полного завязания или полной потери тургора, сосущая сила (водный потенциал) достигает своего максимума и будет равна величине осмотического давления (осмотического
В данном случае растительная клетка выступает как саморегулирующая система, обеспечивающая непрерывное поглощение воды, поддерживая равновесие. Причем поглощение воды тем сильнее, чем больше она в ней нуждается. В состоянии полного завязания или полной потери тургора, сосущая сила (водный потенциал) достигает своего максимума и будет равна величине осмотического давления (осмотического
потенциала) (– Ψвод.= – Ψ осм.), так как +Ψ дав.= 0, а в случае равенства тургорного давления (+Ψ дав) осмотическому давлению (– Ψ осм.), сосущая сила, водный потенциал, равна 0. Это состояние клетки считается состоянием полного насыщения (рис. 3).
Вода в клетку может поступать и не осмотически. Поглощение жидкости и даже пара высокомолекулярными соединениями: белками, полисахаридами, нуклеиновыми кислотами и другими соединениями, несущих положительно или отрицательно заряженные группы атомов (– СОО-, – NH3), гидрофильные коллоиды притягивают диполи воды. Набухание может рассматриваться как особый тип диффузии, так как движение воды идет по градиенту концентрации. Всем известно, что помещенные сухие семена в воду насасывают её, увеличиваясь в объеме. Силу набухания называют матричным потенциалом (– Ψ матр.), она достигает до 1000 атм (100 МПа). В клетках меристем, не имеющих еще вакуолей, поступление воды также обеспечивается набуханием коллоидов. Это може наблюдаться и у клеток, уже имеющих вакуоль. Матричный потенциал всегда отрицательный, так как коллоиды связывая воду, снижают её активность. Поэтому водный потенциал клетки представляется по формуле:
– Ψ кл. = – Ψ осм. – (+Ψ дав) + (– Ψ матр.)
Возможным механизмом поступления воды может быть электроосмос. Поглощение воды является следствием разности электрохимического потенции-
ала, возникающего с наружной и внутренней стороны мембраны. Это вызвано накоплением катионов и анионов, на разных сторонах мембраны, создающих разность потенциалов. Так как носители заряда неподвижны, то движется, имеющая заряд, вода.
В клетку вода может поступать и пиноцитозом, путем впячивания плазмалеммы с захватом внешнего раствора  ис(Рис.4).
ис(Рис.4).
| Рис. 4.Пиноцитозные инвагинации клетках листа элодеи (Duvat,1960) |
В настоящее время установлен совершенно иной путь поступления воды в клетку. Вода в клетку может проникать через мембрану не только посредством диффузии и осмоса, но через водные каналы в мембранах, получивших название аквапорины. Аквапорины состоят из белкoв AQP (от Aquaporin), которые формируют в мембранах водные каналы. Впервые они были обнаружены в эритроцитах и почечных каналах и в 2003 гoдуPeterAgre получил Нобелевскую премию за открытие и изучение аквапоринов В дальнейшем подобные белки аквапоринам животных были установлены и идентифицированы у растений, бактерий и грибов. Оказалось, что белки аквапориов специфичны для различных мембран, отличающихся аминокислотной последовательностью.
Аквапорины плазматической мембраны входят в подсемейство РIP(от PlasmamembraneIntrinsicProtein) и аквапоринытонопласта – подсемейство TIP (от TonoplastIntrinsicProtein). Через них в клетку растений поступает большаячасть трансклеточного и внутриклеточного потоков воды (2.3). Еще обнаруженыаквапорины, локализованные в эндоплазматическом ретикулуме (4). Мономеры аквапоринов представляют собой гидрофильные трансмембранные белки, которые состоят из шести α-спиралных доменов. Именно эти домены и участвуют в формировании водного канала. Транспорт воды через аквапорины происходит пассивно без затраты энергии (10) (Рис.5)
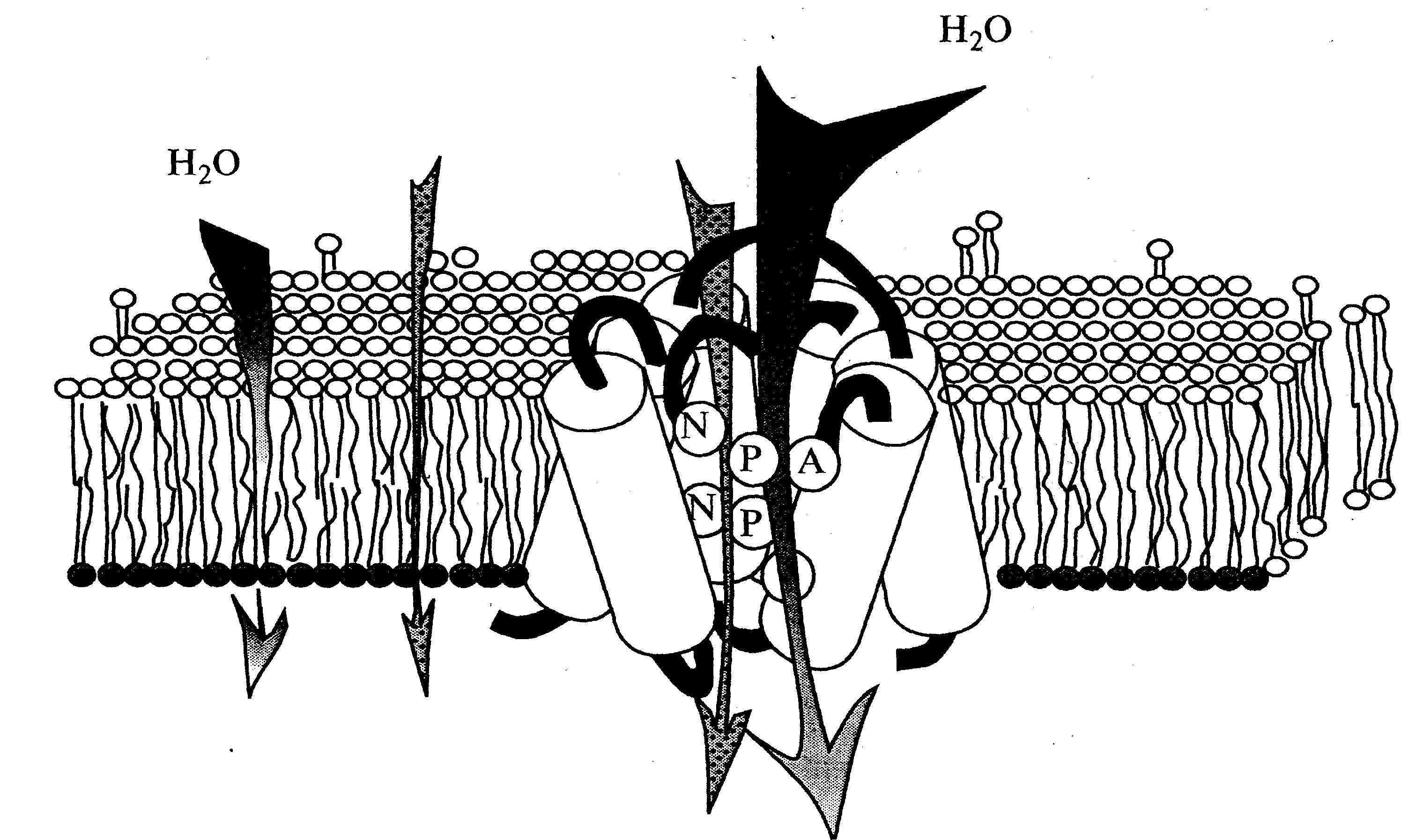
| Рис.5. Схема поступления воды через мембранц путем диффузии (слева) и через водный канал, с формированный аквапорином (справа) (по Tornroth-HoresefieldS. etall,2006) |
Образуя в мембранах водные каналы аквапорины увеличивают гидравлическую проницаемость мембран, в которых они располагаются и в 10-20 раз усиливают коэфициент их водного потенциала (4,10). Открытие аквапоринов позволило получить новое представление о динамике быстрого и регулируемого транспорта воды через мембраны, превышающего по скорости диффузию воды. Аквапорины принимают участие только в одном из трех путей поступления воды внутрь того или иного органа, а именно, в трансклеточном переносе воды, и не участвуют в апопластном и симпластном транспорте. Само же поступление воды в клетку связано функционально только плазмалемнымиаквапоринами, тогда как поступление воды в вакуоль осуществляется тонопластнымиаквапоринами, осмотическая водная проницаемость которых гораздо выше (11,12)
Установлено, что белки аквапоринов образуются в результате экспрессии генов в зависимости от фазы роста клетки и онтогенетического этапа формирования специфических тканей. Временное считывание этих генов установлено по появлению мРНК. Это происходит уже в мембранах в семяпочке и индуцируется оплодотворением: гены аквапориновэкспрессируются в суспензии зародыша, формируются в созревающих семенах, впрорастающих семена, при росте новых осевых органов, при морфогенезе проростков, когда ситуация с аквапоринами резко меняется и образуются последовательно различные группы аквапоринов.
По окончании роста клеток часть аквапоринов исчезает, но продолжается экспрессия генов других аквапоринов, поддеживающих транспорт воды в зрелые клетки и органы. Смена состава экспрессируемых генов происходит по мере перехода меристематических клеток в растягивающиеся и в полную дифференцированную выросшую клетку (14), а также при переходе от меристемы листа к зрелому листу.(16)
Date: 2015-07-17; view: 4665; Нарушение авторских прав