Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Регуляция экспрессии генов про- и эукариот. Теория оперона. Регуляция экспрессии генов у прокариот.Теория Оперона
|
|
Регуляция
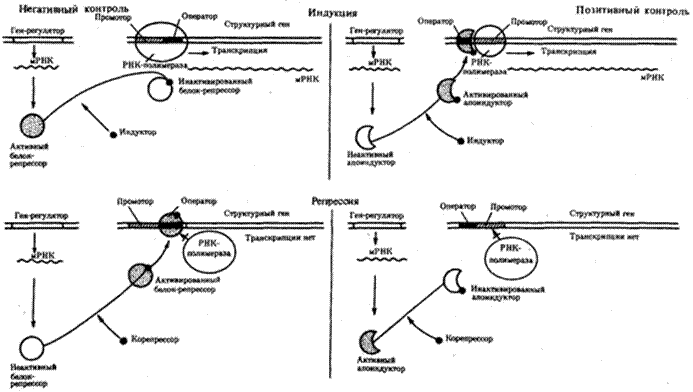
Оперон — это тесно связанная последовательность структурных генов, определяющих синтез группы белков, которые участвуют в одной цепи биохимических преобразований. Например, это могут быть гены, которые детерминируют синтез ферментов, участвующих в метаболизме какого-либо вещества или в синтезе какого-то компонента клетки. Оперонная модель регуляции экспрессии генов предполагает наличие единой системы регуляции у таких объединенных в один оперон структурных генов, имеющих общий промотор и оператор.
Примером участия генетических и негенетических факторов в регуляции экспрессии генов у прокариот может служить функционирование лактозного оперона у кишечной палочки. При отсутствии в среде, на которой выращиваются бактерии, сахара лактозы активный белок-репрессор, синтезируемый геном-регулятором (I), взаимодействует с оператором (О), препятствуя соединению РНК-полимеразы с промотором (Р) и транскрипции структурных генов Z, Y, А. Появление в среде лактозы инактивирует репрессор, он не соединяется с оператором, РНК-полимераза взаимодействует с промотором и осуществляет транскрипцию полицистронной мРНК. Последняя обеспечивает синтез сразу всех ферментов, участвующих в метаболизме лактозы. Уменьшение содержания лактозы в результате ее ферментативного расщепления приводит к восстановлению способности репрессора соединяться с оператором и прекращению транскрипции генов Z, Y, А.

Регуляция активности генов у эукариот.
У человека 25 000 генов
Гены составляют около 10% от ДНК
Кодирующие последовательности составляют около 3% от ДНК
К некодируемым участкам относят: интроны, регуляторные участки гена (промоторы и терминаторы) энхансеры и сайленсоры, повторы в ДНК
Все гены делятся на:
1. репрессированные (больше 90%) т.е в данном типе клеток не транскибируются
2. дерепрессированные могут транскибироваться в данном типе клеток 5-10%
Дерепрессированные делятся на
1. гены конституативные – работают в клетке постоянно, как правило они контролируют жизнено важные ф-ции: ДНК-полимераза, РНК-полимераза, ферменты синтеза АТФ и т.д. Активность генов регулируется способностью РНК-полимеразы распознавать промотр и взаимодействовать с ним.
2. Гены регулируемые - т.е их активность контролируется дополнительными факторами.
2.1 Специфически регулируемая – направлена либо на 1 ген, либо на группу генов:
А) Факторы транскрипции – специальные белки которые контролируют способность РНК-полимеразы распознавать и связываться с промотором. Часто обладают тканевой специфичность т.е в клетках разных тканей факторы транскрипции могут являться разные белки
Рассмотрим действие фактора транскрипции на примере трийодтиронина (Т3). В клетке имеется рецептор Т3. Он связывается с рецептором ретиноевых кислот. Этот комплекс присоединяется к ДНК. К белковому комплексу присоединяется белок ко-репрессор. В таком состоянии ген не активен.
В клетку поступает Т3, проникает в ядро и связывается своим рецептором. В результате изменения комформации (пространственного строения) комплекс рецептора Т3 + рецептор ретиноевых кислот
Отщипывается ко-репрессор и присоединяется белок ко-активатор. В этом случае РНК-полимераза способра распознать промотр гена.
Б) энхансеры – регуляторные участки ДНК, которые усиливают транскирипцию гена
Свойства:
Энхансеры могут находиться как внутри гена так и вне гена.
Один энхансер может стимулировать транскрипцию нескольких генов
Активность энхансера контролируется различными факторами
В) Сайленсоры – регулируемые участки ДНК, которые ингибируют (замедляют) транскрипцию гена. С-ва такие же как и у энхансеров
Г) метилирование азотистых оснований нуклеотидов – высокий уровень метилирования сопровождается блокированием активности гена.
2.2 Регуляция неспецифическая – направлена на большую группу генов:
А) формирование факультативного гетерохроматина на определенном участке ДНК. Гетерохроматин представляет собой очень плотноупакованную ДНК. Транскрипция на этом участке невозможна.
Примеры: у женщин одна Х-хр находится в состоянии гетерохроматина. Таким образом по генам, находящимся в Х-хр женщинский организм формально диплоиден, а фактически гаплоиден.
Мучной червь – у самцов все отцовские хромосомы находятся в состоянии гетерохроматина
18.Особенности хромосомной организации наследственного материала в зависимости от фазы пролиферативного цикла (хроматин, метафазная хромосома). Нуклеосомная модель строения хромосом.
В интерфазе при световой микроскопии он выявляется в виде глыбок, рассеянных в нуклеоплазме ядра. При переходе клетки к митозу, особенно в метафазе, хроматин приобретает вид хорошо различимых отдельных интенсивно окрашенных телец — хромосом.
Диаметр нитчатой структуры интерфазного хроматина равен 100—200, а диаметр одной из сестринских хроматид метафазной хромосомы — 500— 600 нм.
Нуклеосомная модель строения хромосом
1. Нуклеосомиая нить. Этот уровень организации хроматина обеспечивается четырьмя видами нуклеосомных гистонов: Н2А, Н2В, НЗ, Н4. Они образуют напоминающие по форме шайбу белковые тела — коры, состоящие из восьми молекул (по две молекулы каждого вида гистонов)
Молекула ДНК комплектируется с белковыми корами, спирально накручиваясь на них. При этом в контакте с каждым кором оказывается участок ДНК, состоящий из 146 пар нуклеотидов (п.н.). Свободные от контакта с белковыми телами участки ДНК называют связующими или линкерными. Они включают от 15 до 100 п.н. (в среднем 60 п.н.) в зависимости от типа клетки.
Отрезок молекулы ДНК длиной около 200 п. н. вместе с белковым кором составляет нуклеосому.

2. Хроматиновая фибрилла. Дальнейшая компактизация нуклеосомной нити обеспечивается пистоном HI, который, соединяясь с линкерной ДНК и двумя соседними белковыми телами, сближает их друг с другом. диаметр 20—30 нм

Интерфазная хромонема. Укладка хроматиновой фибриллы в петли. В их образовании, принимают участие негистоновые белки, которые способны узнавать специфические нуклеотидные последовательности вненуклеосомной ДНК, отдаленные друг от друга на расстояние в несколько тысяч пар нуклеотидов. Эти белки сближают указанные участки с образованием петель из расположенных между ними фрагментов хроматиновой фибриллы Хроматиновая фибрилла диаметром 20—30 нм преобразуется в структуру диаметром 100—200 нм. Отдельные участки интерфазной хромонемы подвергаются дальнейшей компактизации, образуя структурные блоки,
3.
4. Метафазная хромосома. Суперкомпактизация хроматина. Отдельные хромосомы становятся хорошо различимы. Этот процесс начинается в профазе, достигая своего максимального выражения в метафазе митоза и анафазе. В телофазе митоза происходит декомпак-тизация вещества хромосом, которое приобретает структуру интерфазного хроматина. Описанная митотическая суперкомпактизация облегчает распределение хромосом к полюсам митотического веретена в анафазе митоза.
Пресинтетический период (2n 2c, где n — число хромосом, с — число молекул ДНК); Синтетический период (2n 4c) Постсинтетический период (2n 4c); Профаза (2n 4c); Метафаза (2n 4c); Анафаза (4n 4c);Телофаза (2n 2c в каждой дочерней клетке).
19.Хромосома, ее химический состав и морфологическая характеристика. Понятие об эухроматине и гетерохроматине.
Строение хромосомы.
Морфологическая характеристика
Каждая хромосома имеет центральную перетяжку (центромера). Вблизи первичной перетяжки имеется место прикрепления нитей веретена деления. 2 плеча. На концах плеч распоалгаются теломеры или теломерные участки. В хромосомах имеются участки эухроматина, факультативного гетерохроматина, конституитивного гетерохроматина. Эти участки не выполняют генетической функции. Они обеспечивают: структурно-функциональную целостность хромосомы, обеспечивают взаимодействие с нитями веретена деления, обеспечивают взаимодействие с оболочкой ядра.
Химический состав хромосомы
Хромосомы состоят в основном из ДНК и белков, которые образуют нуклеопротеиновый комплекс—хроматин
Белки составляют значительную часть вещества хромосом. На их долю приходится около 65% массы этих структур. Все хромосомные белки разделяются на две группы: гистоны и негистоновые белки.
Гистоны представлены пятью фракциями: HI, Н2А, Н2В, НЗ, Н4. Являясь положительно заряженными основными белками, они достаточно прочно соединяются с молекулами ДНК, чем препятствуют считыванию заключенной в ней биологической информации. В этом состоит их регуляторная роль. Кроме того, эти белки выполняют структурную функцию, обеспечивая пространственную организацию ДНК в хромосомах
Число фракций негистоновых белков превышает 100. Среди них ферменты синтеза и процессинга РНК, редупликации и репарации ДНК. Кислые белки хромосом выполняют также структурную и регуляторную роль. Помимо ДНК и белков в составе хромосом обнаруживаются также РНК, липиды, полисахариды, ионы металлов.
РНК хромосом представлена отчасти продуктами транскрипции, еще не покинувшими место синтеза. Некоторым фракциям свойственна регуляторная функция.
Массовые соотношения ДНК: гистоны: негистоновые белки: РНК: липиды — равны 1:1:(0,2—0,5):(0,1—0,15):(0,01—-0,03). Другие компоненты встречаются в незначительном количестве.
Функции: 1) хранение наследственной инфо 2) использование наследствен инфо для создания и поддержания клеточной орг-ции. 3)регуляция считывания наследств инфо 4)размножение н.и. путем редупликации
5)передача н.и.
Структура хромосом:
1)первичная структура-это нуклеосомная нить. Гистоны Н2а, Н2в Н3 Н4 образуют КОР (по 2 молекулы каждой фракции=>8 молекул)
На КОР наматывается ДНК длиной 146 пар нуклеотидов. КОР+ДНК=нуклеосома
Через 40 пар нуклеотидов Днк образуется следующая неклеосома. Та ДНК которая не связана с кором называется линкерной. 2)вторичная структура-нуклеопротеидная фибрила (гистон Н1)
За счет гистонов Н1 происходит сближение двух соседних нуклеосом. При этом линкерная Днк образует петлю. Молекулы Н1 находятся под линкерными. 3) Третичная структура-интерфазная хромонема. За счет негистоновых белков происходит упаковка в петли нуклеопротеидной фибриллы. 4)четвертичная структура-метафазная хроматида.
Дальнейшая спирализация за счет ионов металлов.
В зависимости от состояния хроматина выделяют эухроматиновые участки хромосом, отличающиеся меньшей плотностью упаковки в неделящихся клетках и потенциально транскрибируемые, и гетерохроматиновые участки, характеризующиеся компактной организацией и генетической инертностью.
Различают конститутивный (структурный) и факультативный гетерохроматин.
Конститутивный гетерохроматин содержится в околоцентромерных и теломерных участках всех хромосом, а также на протяжении некоторых внутренних фрагментов отдельных хромосомОн образован только нетранскрибируемой ДНК. Вероятно, его роль заключается в поддержании общей структуры ядра, прикреплении хроматина к ядерной оболочке, взаимном узнавании гомологичных хромосом в мейозе, разделении соседних структурных генов, участии в процессах регуляции их активности.
Примером факультативного гетерохроматина Х-хр у женщин. Гены этой хромосомы не транскрибируются. Образование факультативного гетерохроматина за счет генетического материала других хромосом сопровождает процесс клеточной дифференцировки и служит механизмом выключения из активной функции групп генов, транскрипция которых не требуется в клетках данной специализации.
20.Хромосомная теория. Карты хромосом (физические, рестрикционные, химические, генные). Принципы составления карт хромосом.
Хромосомная теория
1. Гены в хоромосомах располагаются линейном порядке. Участок, который занимает 1 ген назыают локусом. В индентичных локусах гомологичных хромосом располагаются гены, которые влияют на формирование одного признака.
2. Гены располагаются в хромосомах, которые в основном находятся в ядре. Исключение: митохондрии, хлоропласты. В свяызи с этим имеется явление цитоплозматической наслественности. В этом случае признак передается только от матери. Это нетрадиционный способ наследования
3. Гены, лежащие в одной хромосоме образуют группу сцепления и поэтому как правило наследуются совместно. Врезультате в ряду поколений отмечается наличие устойчивого комплекса признаков. Например, у человек У-хр содержит:
1. Ген-контроль
2. Ген-мутация, в котором вызывается формирование перепонок между пальцами ног
3. Ген-мутация, в котором вызывается повышенное оволосенение
4. Количество групп сцепления у гомогаметного пола равно гаплоидному числу хромосом. Учеловека гомогаметный пол женский. Поэтому количество групп сцепления равно 23. У гетерогаметного пола число групп сцепления = n+1. У человека гетерогаметный пол мужской. Поэтому кол-во групп сцепления у него 24
5. Сцепление генов неабсолютно и может нарушаться при кроссинговере, который происходит в профазе 1 мейоза. Сила сцепления между генами зависит от расстояния между ними – чем больше расстояние, тем меньше сила сцепления и выше частота кроссинговера. Расстояние между генами измеряют в морганидах (1 морг = 1% кроссинговера)
6. Каждый биологический вид в своих диплоидных клетках содержит уникальный набор хромосмо – кареотип. Кариотип различкается по количеству хромосом, их размерам или строению.
Карты хромосом
Генетические карты хромосом — это схема взаимного расположения и относительных расстояний между генами определенных хромосом, находящихся в одной группе сцепления.
Физическая карта – графическое представление порядка следования физических маркеров (фрагментов молекулы ДНК), расстояние между которыми определяется в парах нуклеотидов. Рестрикционная карта – вид физической карты, на которой указан порядок следования и расстояния между сайтами расщепления ДНК рестриктазами (обычно участок узнавания рестриктазы 4-6 п.н.). Маркерами этой карты являются рестрикционные фрагменты/сайты рестрикции.
Химическая карта – состав.
Для составления карт хромосом рассчитывают взаимное расстояние между отдельными парами генов и затем определяют расположение этих генов относительно друг друга. Так, например, если три гена расположены в следующем порядке: А В С, то расстояние между генами А и С (процент рекомбинаций) будет равно сумме расстояний (процентов рекомбинаций) между парами генов АВ и ВС. Если гены расположены в порядке: А С В, то расстояние между генами А и С будет равно разности расстояний между парами генов АВ и СВ.
Date: 2016-07-22; view: 2572; Нарушение авторских прав