Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Физиологическая основа
|
|
Мышца представляет собой массу ткани, состоящей из миллионов отдельных мышечных волокон, соединенных вместе и работающих согласованно. Каждое мышечное волокно — это тонкая нить толщиной всего лишь около 0,1 мм и до 300 мм длиной. При стимуляции электрическим потенциалом действия это волокно сокращается, иногда примерно до половины первоначальной длины. Мышечные волокна функционально связаны в двигательные единицы. Число волокон в каждой такой единице зависит от типа мышцы. Мышцы, участвующие в тонких двигательных коррекциях (как, например, при фиксации объекта глазами), могут иметь в каждой единице всего по 10 волокон, тогда как в мышцах, осуществляющих более грубую регулировку при поддержании позы, в одной двигательной единице может быть до 3000 мышечных волокон. Мышца, таким образом, состоит из группы двигательных единиц.
На рис. 8.2 показано анатомическое строение простой двигательной единицы. Каждая единица управляется одним мотонейроном (двигательной нервной клеткой), расположенным в спинном мозгу (мотонейроны для мышц тела) или в стволе головного мозга (для мышц головы). Разветвления аксона нервной клетки подходят к ряду мышечных волокон.
Сокращение мышц происходит в результате совместного действия большого числа двигательных единиц. Общая величина мышечного сокращения зависит от числа активированных двигательных единиц и частоты получаемых ими нервных импульсов. Обычно импульсы рассредоточены во времени, так что плавное движение получается в результате разновременного сокращения двигательных единиц. При некоторых

Рис. 8.2. Двигательная единица.
заболеваниях двигательные единицы начинают работать почти синхронно, что приводит к тремору или подергиваниям.
Поверхностная ЭМГ суммарно отражает разряды двигательных единиц, вызывающие сокращение. Поскольку ее регистрируют с поверхности кожи, на самом деле все обстоит несколько сложнее. Разряды единиц, располагающихся на разной глубине от поверхности, могут ослабляться в неодинаковой степени. Но, как мы уже говорили, общий уровень электрической активности хорошо соответствует общей величине развиваемого мышечного напряжения.
Такое соответствие будет наилучшим, если электроды аккуратно разместить над данной группой мышц. На рис. 8.3 изображены главные мышечные группы тела. Чаще всего, вероятно, регистрируют активность трапецевидной мышцы (на задней стороне шеи) и плече-лучевой (на предплечье). Как мы уже отмечали, для исследования выбирают те мышечные группы, которые должны лучше всего отражать тип активности, ожидаемый при выполнении данного экспериментального задания. Затем можно установить в опыте, как выглядит картина активации этих групп мышц.
Эмоции и активация (arousal)
Не так давно психофизиологи снова вернулись к изучению активности лицевой мускулатуры при эмоциональных состояниях. Больше ста лет назад Чарлз Дарвин (Darwin, 1872)
Глава
Мышцы
!■
|
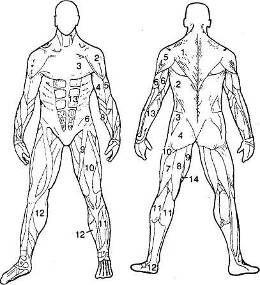
Рис. 8.3. Важнейшие мышцы человека.
Слева (вид спереди): /— m. sterno-cleido-mastoideus; 2 — т. deltoides; 3 — т. pectoralis maior; 4 — т. biceps brachii; 5 — т. triceps brachii; 6 — т. obliquus abdominis externus; 7 — m. brachiora-dialis; S — m. flexor carpi ulnaris; 9 — m. sartorius; 10 — m. quadriceps femoris; // — m. tibialis anterior; 12 — m. gastrocnemius; 13 — m. rectus abdominis.
Справа (вид сзади): / — т. trapezius; 2 — т. latissimus dorsi; 3 — т. gluteus medjus; 4 — т. gluteus maximus; 5 — m. deltoides; 6 —m. triceps brachii; 7 — m. biceps femoris; 8 — m. semitendinosus; 9 — m. gracilis; 10 — m. vastus lateralis; // — m. gastrocnemius; 12 — ахиллово сухожилие; 13 — т. flexor carpi radialis; 14 — m. semimem-branosus.
в своей книге «Выражение эмоций у человека и животных» выск-азал мысль, что выражение лица — это один из объективных индикаторов нашей богатой эмоциональной жизни. В частности, он утверждал, что различные выражения лица самим устройством организма предназначены для отображения разных чувств. Разгневанный банту очень похож на разгневанного жителя Нью-Йорка. Позже было проведено много психологических исследований, в которых испытуемые должны были находить соответствие между различными выражениями лица %и определенными чувствами (Woodworth, Schlosberg, 1954). Подводя итоги работам последних лет, Экман (Ekmann, 1971) пришел к выводу, что у людей, принадлежащих к разным культурам, существует единообразие в выражении по крайней
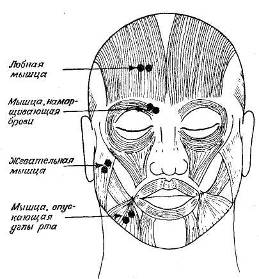
Рис. 8.4. Мышцы лица и положение электродов при записи активности четырех
главных мышц. (Рисунок предоставлен Гэри Э. Шварцем.)
мере следующих основных эмоций: радости, печали, гнева, страха, удивления и отвращения.
В последние годы в Бостонском психиатрическом центре Эриха Линдемана была начата работа по соотнесению выражений лица с электрической активностью лицевой мускулатуры (Schwartz et al., 1976). На рис. 8.4 показаны группы лицевых мышц, а также места расположения электродов при этих исследованиях.
В этих экспериментах испытуемые, спокойно сидевшие в затемненной комнате, должны были представлять себе разного рода события. В первых работах было обнаружено, что у испытуемых, которых, как детей в «Питере Пэне», просили думать о чем-нибудь радостном, усиливается электрическая активность в мышцах, опускающих углы рта, и соответственно снижается активность в мышце, наморщивающей брови. Когда затем те же испытуемые представляли себе печальные картины, у них усиливалась активность в мышце, наморщивающей брови, и в лобной мышце. Эти изменения предшествуют внешнему изменению выражения лица и позволяют поэтому более тонко различать выражения, чем это возможно при простом наблюдении. Таким образом, предварительные данные показали, что запись ЭМГ, по-видимому," отражает весьма малые изменения в мышцах, которые нельзя заметить невооруженным глазом.
5 Зак. 699
Глава 8
 В дальнейшем при работе с больными, страдающими депрессией, те же исследователи обнаружили, что, по крайней мере судя по реакции ЭМГ, этим больным труднее воспроизводить «счастливые» мысли, чем испытуемым контрольной группы. В то же время представления, вызывающие печаль или гнев, были у них такими же яркими, как у всех остальных людей. Наконец, когда обе группы должны были представить себе «обычный день», то у здоровых людей ЭМГ была сходна с ЭМГ при радостных мыслях, тогда как у больных с депрессией она была больше похожа на картину, наблюдаемую при печальных мыслях.
В дальнейшем при работе с больными, страдающими депрессией, те же исследователи обнаружили, что, по крайней мере судя по реакции ЭМГ, этим больным труднее воспроизводить «счастливые» мысли, чем испытуемым контрольной группы. В то же время представления, вызывающие печаль или гнев, были у них такими же яркими, как у всех остальных людей. Наконец, когда обе группы должны были представить себе «обычный день», то у здоровых людей ЭМГ была сходна с ЭМГ при радостных мыслях, тогда как у больных с депрессией она была больше похожа на картину, наблюдаемую при печальных мыслях.
Подобные эксперименты демонстрируют способность ЭМГ отвечать на весьма тонкие изменения. Отметим также, что эти исследования акцентируют общую картину (паттерн) мышечной активности. Выражение лица определяется не какой-то отдельной мышечной группой, а совместным действием многих мышц.
Головной мозг
В некотором смысле психофизиология — это изучение мозга. Если у человека в ответ на неприличное слово изменяется электропроводность кожи, то это следствие того, что мозг классифицирует данное слово как неприличное и соответственно определяет реакцию симпатической нервной системы. Когда у свидетеля при перекрестном допросе учащается мигание, то это мозг классифицирует данную ситуацию как напряженную и соответствующим образом реагирует. Для каждой ситуации мозг формирует какой-то уникальный комплекс физиологических реакций. Мозг — это ведущий орган, каждый человек — это его мозг.
Теперь мы умеем регистрировать активность мозга в нейтральном состоянии. Поскольку мозг — орган очень сложный и функционирование его наименее понятно для нас, его электрическая активность очень сложна.
История вопроса
Многие из наших современных представлений о функциях мозга восходят к Галлю — отцу френологии. В начале XIX столетия Галль провозгласил, что мозг человека состоит из двадцати семи отдельных органов, каждый из которых ответствен за одну из наших основных способностей. Далее он утверждал, что, исследовав распределение шишек на черепе, можно видеть, насколько хорошо развит каждый из этих органов, и таким образом предсказать черты личности данного человека. Можно было, например, ожидать, что обладатель шишки на левом виске будет склонен к стяжательству. В свое время Галля осудила религия. Сегодня его теорию отвергает наука. Форма черепа не соответствует форме мозга, и вся система классификации способностей не выдерживает критики.
Однако Галль заронил зерно мысли о локализации функций в мозгу. Хотя при нашем уровне знаний уже невозможно представлять себе, что каждая отдельная область мозга управляет определенным типом поведения, мы, бесспорно, можем говорить по крайней мере об известной степени локализации мозговых функций. Например, анатомические связи, идущие от глаз, образуют четко обособленный путь от сетчатки 5*
Глава 9
Головной мозг
к области в затылочной коре. Одна из главных задач современных исследований мозга — попытаться выяснить, в какой степени локализованы функции различных типов.
В 1875 году английский хирург Ричард Кэйтон впервые показал, что у животного можно зарегистрировать электрическую активность мозга. Кэйтон, так же как и Галль, стремился доказать, что разные отделы мозга выполняют различные функции. При помощи аппаратуры, которая по современным стандартам была чрезвычайно малочувствительной, Кэйтону удалось показать, что на поверхности сенсорной коры кролика при воздействии света на глаза возникали характерные электрические изменения. Он отметил также, что в отсутствие света у кролика можно наблюдать регулярную фоновую электрическую активность.
Прошло более пятидесяти лет, прежде чем сходные наблюдения были сделаны на человеке. Австрийский психиатр Ханс Бергер (Berger, 1929) обнаружил, что с поверхности черепа можно зарегистрировать «мозговые волны». В серии экспериментов, проведенных во время хирургических операций на мозге, Бергер смог показать, что некоторая часть этих электрических потенциалов действительно принадлежит мозгу, а не обусловлена просто активностью мышц головы. Кроме того, он установил, что электрические характеристики этих сигналов зависят от состояния испытуемого. Наиболее заметными были синхронные волны относительно большой амплитуды (около 50 микровольт) с характерной частотой около 10 циклов в секунду. Бергер назвал их «альфа-волнами» и противопоставил высокочастотным «бета-волнам», которые появляются, когда человек переходит в более активное состояние.
В качестве электродов Бергер использовал довольно большие подушечки, пропитанные солевым раствором, которые прикреплялись ко лбу и к затылку. Он считал, что обнаружил физиологический показатель, аналогичный электрокардиограмме: подобно тому как ЭКГ может быть индикатором общего состояния сердечной мышцы, электроэнцефалограмма (ЭЭГ) представляет собой индикатор общей активности мозга.
Позднейшие исследователи показали, что ЭЭГ качественно отличается от открытых ранее более простых показателей активности вегетативной нервной системы. Периодические сокращения сердца и связанные с ними сдвиги потенциала — это сама простота по сравнению с ужасающей сложностью ЭЭГ. Интуитивно мы могли предполагать, что код работы „мозга бесконечно более сложен, чем законы сокращения мышц. ЭЭГ эти ожидания оправдывает, и даже с избытком. Для. интерпретации наблюдаемых волн существенно не только место их возникновения: сложность их формы как будто бросает
вызов исследователям, пытающимся найти в них хоть какой-то смысл.
Современники Бергера отнеслись к его сообщению скептически, и «мозговые волны» стали общепризнанным фактом только после того, как Эдриан и Мэттьюз осуществили наглядную демонстрацию записи ЭЭГ на заседании Английского физиологического общества в 1935 году. К аппарату подсоединили самого д-ра Эдриана, и он показал, как при открывании глаз подавляется высокоамплитудный альфа-ритм.
Последующие годы были волнующими для исследователей мозговых потенциалов. И хотя работы сдерживались недостатком средств в период экономической депрессии, ученые с энтузиазмом проникали в тайны глубин мозга, пользуясь крупными успехами электронной усилительной техники и применяя ее с необычайной изобретательностью. В то время в статьях по электроэнцефалографии электронные схемы встречались так же часто, как и записи ЭЭГ.
Предварительные данные Бергера о четких электроэнцефалографических коррелятах эпилепсии были подтверждены Гиббсом и сотр. (Gibbs et al., 1935). В том же году Корнмюллер убедительно показал, что при записи с разных участков черепа регистрируются колебания разной формы. В 1936 году Уолтер обнаружил, что при опухолях мозга в окружающей их ткани обычно появляются аномальные медленные волны и что ЭЭГ можно использовать для уточнения локализации таких опухолей.
После перерыва, связанного со второй мировой войной, исследования ЭЭГ продолжали расширяться. В 1949 году вышел первый номер Electroencephalography and Clinical Neurophysiology — международного журнала, целиком посвященного изучению электрической активности мозга.
В последующий период не все шло гладко. Возможность регистрировать ЭЭГ у бодрствующих людей вселила в ученых большой оптимизм: казалось, открывается путь к пониманию взаимоотношений между «душой» и телом. Некоторые даже предвидели день, когда мы сможем «читать» то, что происходит в сознании, просто рассматривая характер мозговых волн. Было начато бесчисленное множество исследований, направленных на поиски электроэнцефалографических коррелятов интеллекта, особенностей личности, поведения. Результаты в большей своей части оказались разочаровывающими.
В первом обзоре англоязычной литературы по ЭЭГ Джаспер (Jasper, 1937) предсказывал: «Попытки установить соответствие между особенностями ЭЭГ и столь же туманными психологическими процессами, такими, как „внимание", „сознание",..мышление" и разного рода сложные атрибуты „личности"...,
Глава 9
Головной мозг
 могут только усугубить путаницу, существующую сейчас в психологической терминологии». С тех пор были проведены тысячи исследований, и они во многом подтвердили это мрачное предсказание, хотя оно не во всем оказалось верным. В мозгу человека больше 10 миллиардов нервных клеток, сплетенных в плотную сеть взаимными связями. Даже в самых тонких записях ЭЭГ неизбежно выявляется лишь слитная трескотня сотен тысяч клеток, приглушенная и искаженная черепом. «Мы подобны слепым, пытающимся понять работу фабрики, прислушиваясь снаружи к ее шуму» (Margerison et al., 1967).
могут только усугубить путаницу, существующую сейчас в психологической терминологии». С тех пор были проведены тысячи исследований, и они во многом подтвердили это мрачное предсказание, хотя оно не во всем оказалось верным. В мозгу человека больше 10 миллиардов нервных клеток, сплетенных в плотную сеть взаимными связями. Даже в самых тонких записях ЭЭГ неизбежно выявляется лишь слитная трескотня сотен тысяч клеток, приглушенная и искаженная черепом. «Мы подобны слепым, пытающимся понять работу фабрики, прислушиваясь снаружи к ее шуму» (Margerison et al., 1967).
Обзор показателей ЭЭГ
Некоторые из наиболее интересных современных работ в области физиологической психологии посвящены регистрации разрядов отдельных нейронов мозга. Такая регистрация одиночных нейронов, как ее называют, требует введения электродов в ткань мозга, и поэтому ее производят обычно у животных.
Изучая функции мозга у человека, психофизиолог в большинстве случаев вынужден отводить электрическую активность от поверхности головы. Только такое отведение мы и будем называть электроэнцефалографией, хотя этот термин часто употребляют для обозначения прямой регистрации с поверхности коры или даже записи с помощью подкорковых вживленных электродов.
Возможно, что в будущем психофизиолог сможет отводить активность от глубоких структур мозга, не прибегая к хирургии. Все электрические токи создают магнитные поля, и токи в нервной системе не составляют исключения. Сложные электрические ритмы мозга генерируют очень слабые магнитные поля. Коэн (Cohen, 1972) в Масса чу сетском технологическом институте исследовал возможность регистрации этих полей. Располагая детекторы примерно на расстоянии 5 см от кожи в сильно экранированной комнате, он смог записывать магнитную активность. Это — немалое достижение. Напряженность таких полей имеет величину порядка 10~9 гаусс, что составляет около 1/100 000 случайного магнитного фона в условиях города. Поэтому магнитоэнцефалография требует в настоящее время очень сложного и дорогого оборудования. Однако благодаря упразднению электродов этот метод, возможно, будет давать такую информацию об электрической активности мозга, которую нельзя получить с помощью обычных методов.
Сегодня ЭЭГ остается наиболее перспективным, но пока наименее расшифрованным источником данных для психофизиолога. Одна из ее самых поразительных черт — это ее
спонтанный, автономный характер. Регулярные электрические осцилляции прекращаются только с наступлением смерти: даже при глубокой коме и наркозе наблюдается особая характерная картина мозговых волн. Большинство исследований ЭЭГ связано с анализом этих спонтанных ритмов.
В конце главы мы рассмотрим более сложные методы такого анализа—усреднение вызванных потенциалов (УВП) и отрицательного отклонения медленного потенциала (ООМП). Оба показателя основаны на «усреднении» спонтанных ритмов. Главный принцип состоит в том, что при многократном повторении какого-то раздражителя низкоамплитудные кратковременные изменения ЭЭГ в ответ на него можно выделить, устранив фоновую электрическую активность.
Но вернемся к анализу спонтанных ритмов. Простейший подход состоит в просмотре записи и в формировании «клинического впечатления» (об этом будет говориться позже). Подобный метод наиболее полезен в случае резких изменений при патологических состояниях, например при эпилепсии. Изучение более тонких изменений ЭЭГ в зависимости от психологического состояния начинается с рассмотрения частот и (или) амплитуд мозговых волн.
Начнем с альфа-ритма. Как вы помните, Бергер (Berger, 1929) дал это название относительно высокоамплитудным синхронным волнам с частотой около 10 Гц, появлявшимся главным образом в затылочных отведениях, когда человек бодрствовал, но находился в расслабленном состоянии. Бергер противопоставлял этот ритм колебаниям, которые он назвал бета-волнами,— низкоамплитудным асинхронным колебаниям более высокой частоты, по-видимому сопровождавшим состояние активного бодрствования.
Когда все эти волны были лучше изучены, альфа-волнами стали называть всю активность с частотой в полосе 8—13 Гц, а бета-волнами — все колебания с доминирующей частотой более 13 Гц. Ранние наблюдения Бергера о связи бета-волн с состоянием относительной активности были в общем подтверждены, по крайней мере на уровне упрощенного рассмотрения. Волны более высокой частоты обычно считают признаком большей активации (Lindsley, 1951).
К бергеровской дихотомической классификации были добавлены две полосы частот — тета (4—8 Гц) и дельта (менее 4 Гц). Эти четыре основные категории колебаний схематически представлены на рис. 9.1. Следует подчеркнуть, что это разбиение на группы по частоте более или менее произвольно — оно не соответствует каким-то физиологическим категориям. Их связь со степенью «психической активации» представляется довольно слабой, так как есть много исклю-
Глава 9
Головной мозг

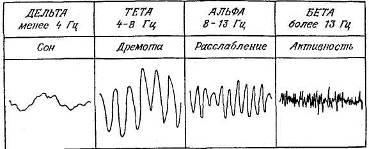
 Рис. 9.1. Классификация мозговых волн по частоте.
Рис. 9.1. Классификация мозговых волн по частоте.
•Указаны приблизительные поведенческие корреляты различных частотных диапазонов, обычно рассматриваемые в теории активации.
чений. Например, у взрослых людей при эмоциональных переживаниях обнаруживаются колебания тета-частоты. Кроме того, эта классификация не подходит одинаково для всех возрастов. Альфа-ритм появляется лишь в раннем подростковом возрасте.
Тем не менее это основные традиционные категории частот мозговых колебаний. В любой данный момент у любого взрослого человека может обнаружиться картина, отличная от описанных выше типов. Для определения доминантной полосы частот нередко используют анализ Фурье, о котором мы будем говорить в этой главе позже.
Другая важная характеристика спонтанных ритмов мозга — это их амплитуда, т. е. величина электрических изменений. Амплитуда и частота колебаний связаны друг с другом. Амплитуда более высокочастотных бета-волн составляет обычно только одну десятую амплитуды более медленных альфа-волн у того же человека.
При первых попытках более систематического анализа изменений ЭЭГ обычно измеряли и частоту, и амплитуду. Одна из наиболее часто используемых зависимых переменных — это «время альфа-ритма», т. е. процент времени, занимаемый альфа-ритмом. Кажущаяся точность этого термина несколько обманчива. Как мы видели, альфа-волны — это просто колебания с частотой 8—13 Гц. однако фактически любые сложные колебания включают и некоторую долю активности в этом диапазоне частот. Таким образом, критерием «наличия альфа-ритма» должна быть какая-то минимальная амплитуда, относительная или абсолютная, таких волн. При «абсолютном ме*тоде» устанавливают некоторую условную границу, например любые колебания с частотой 8—13 Гц и амплитудой 30 микровольт рассматривают как альфа-ритм, тогда как при
наличии колебаний той же частоты, но с амплитудой 29,9 микровольта и ниже считают, что альфа-активность отсутствует. При «относительном методе» эта условная граница определяется для каждого человека отдельно. Обычно перед началом эксперимента испытуемому дают пять минут отдыха. В этот период определяют среднюю амплитуду альфа-волн. Это может быть 60 микровольт у одного испытуемого и 45 у другого. Считают, что альфа-активность имеется во всех случаях, когда активность с частотой 8—13 Гц превышает некоторую, заранее условленную долю (например, 1/3) найденной средней величины. Так, в нашем примере о первом испытуемом можно было бы сказать, что у него имеется альфа-активность, когда амплитуда колебаний с частотой 8—13 Гц превышает у него 20 микровольт, тогда как для второго в том же исследовании условной границей будет амплитуда 15 микровольт. Эта методика представляет собой первый шаг в компенсации индивидуальных различий для выделения разных реакций при решении данной задачи. Ясно, что она менее пригодна для изучения различий между испытуемыми. Сейчас исследователи ЭЭГ пытаются измерять несколько частотных полос одновременно и рассматривать всю электрическую активность в целом.
Еще один традиционный показатель ЭЭГ — это «блокада альфа-ритма», т. е. внезапное очень резкое уменьшение амплитуды альфа-волн, которое обычно происходит при предъявлении раздражителей. Например, если на глаза испытуемого падает свет, в затылочных отделах мозга у него обычно происходит блокада альфа-ритма. Как и в случае «времени альфа-ритма», точный критерий блокады может быть разным у разных исследователей. Следует также отметить, что при блокаде альфа-ритма спонтанные ритмы мозга не исчезают: происходит лишь замена активности с частотой 8—13 Гц более высокими или более низкими частотами.
Таким образом, хотя частотный и амплитудный анализ ЭЭГ представляет собой шаг в направлении большей точности, он еще далеко не удовлетворяет нас в этом отношении. Прежде чем рассматривать более точные (но и более дорогостоящие) методы анализа, нам нужно будет рассмотреть физиологическую основу ЭЭГ.
Физиологическая основа ЭЭГ
До сих пор в электроэнцефалографии преобладает эмпирическое направление — занимаются в основном сравнением ЭЭГ у разных людей и при разных психологических состояниях. Огромное большинство исследователей лишь поверхностно
Глава 9
Головной мозг
 интересуется физиологическими основами ЭЭГ. Главное внимание уделялось в этой области причинам синхронности альфа-ритма. Поскольку обычная ЭЭГ отражает суммарную активность сотен тысяч клеток, в предлагаемых объяснениях подчеркивалась роль механизмов, обеспечивающих согласованную активность больших клеточных масс.
интересуется физиологическими основами ЭЭГ. Главное внимание уделялось в этой области причинам синхронности альфа-ритма. Поскольку обычная ЭЭГ отражает суммарную активность сотен тысяч клеток, в предлагаемых объяснениях подчеркивалась роль механизмов, обеспечивающих согласованную активность больших клеточных масс.
Прежде чем перейти к этим гипотезам, отметим, что некоторые скептики утверждали, будто альфа-ритм совсем или почти не связан с истинной корковой активностью, что это просто артефакт. Кеннеди (Kennedy, 1959) полагал, что альфа-ритм обусловлен механическими пульсациями геля (в данном случае мозга), которые в свою очередь вызывают пульсации электрического потенциала. Иными словами, альфа-ритм возникает из-за того, что мозг в черепе дрожит. Регистрация альфа-ритма от глубинных структур мозга (Andersen, Andersson, 1968), по-видимому, опровергает эту теорию.
К такой же категории относятся представления Липпольда и Новотны (Lippold, Novotny, 1970). Эти авторы провели серию изящных экспериментов, результаты которых говорят в пользу того, что затылочный альфа-ритм — это просто отражение колебаний роговично-сетчаточного потенциала, связанных с тремором наружных мышц глаза. В ответ на это Шоу и сотр. (Shaw et al., 1970) привели пример ЭЭГ 27-летней женщины, где был виден типичный альфа-ритм, исчезавший при решении в уме арифметических задач, хотя у нее из-за осложнений двусторонней глаукомы были удалены оба глаза. Против того, что движения глаз — единственная основа альфа-ритма, говорит тот факт, что при изучении механизмов генерации альфа-ритма обычно используют кураризированных животных, у которых наружные мышцы глаз парализованы. Выдвигался также ряд более серьезных физиологических гипотез относительно центральных механизмов, которые могли бы лежать в основе альфа-активности. Линдсли (Lindsley, 1956) утверждал, что происходит синхронизация случайных изменений возбудимости отдельных нейронов. Бард (Bard, 1961) связывал альфа-ритм с метаболическими процессами в клетках. Экклс (Eccles, 1953) предполагал, что за альфа-ритм ответственны реверберирующие цепи нейронов в коре. По-видимому, наиболее обоснованным представляется мнение Андерсена и Андерсона (Andersen, Andersson, 1968). В результате тщательного рассмотрения нейрофизиологической литературы они пришли к выводу:
* «Эксперименты, в которых после удаления коры сохранялась ритмическая активность таламуса, опыты с деафферентацией путем изоляции коры в острых и хронических условиях, результаты поверхностного или общего охлаждения мозга, локального воздействия фармакологическими препаратами — все указывает на то, что таламическая ритмическая активность служит существенно
важным генератором ритма. Это не означает, что кора сама не способна к генер-ации ритмической активности, однако в таламусе тенденция к ритмичности настолько сильнее, что в норме эта структура выступает как водитель ритма».
Эти исследователи предполагают, что разные участки таламуса индуцируют альфа-ритм в разных областях коры. В психофизиологии намечается сейчас больший интерес к психологической интерпретации картин ЭЭГ, получаемых при отведении от того или иного участка.
Локализация функций
Первоначальные наблюдения Бергера (Berger, 1929) базировались на убеждении, что ЭЭГ отражает общую активность мозга. Психофизиологи в общем склонны были разделять это мнение. Отсюда следовало, что, если электроды расположены в области черепа, точное положение их не столь существенно. А это в свою очередь подразумевает «холистическую» точку зрения на функции мозга в противоположность представлению о специфической локализации функций.
Когда Галль выдвинул свою систему френологии, она оказалась, как сказал Эдмунд Дж. Боринг в своей «Истории экспериментальной психологии» (Boring, 1950), «примером теории, которая, будучи в основном ошибочной, была верной ровно настолько, чтобы способствовать прогрессу научной мысли». Ибо Галль не только пробудил интерес к анатомии коры мозга, но воскресил также идею о локализации функций и распространил ее на сложные психические процессы.
За полтора столетия, прошедшие с тех пор, представление о локализации функций несколько раз входило в фавор и впадало в немилость. Исследования Пьера Флуранса в начале XIX века говорили в пользу холистического взгляда. На основе наблюдений, которые оказались ошибочными, он сделал вывод о диффузном представительстве сложных психических функций в мозгу. Эта концепция «эквипотенциальности» различных участков мозга властвовала в западной медицине вплоть до 1861 года, когда Поль Брока на заседании Парижского антропологического общества представил результаты своих наблюдений над больными с повреждениями мозга. Он продемонстрировал препараты мозга нескольких больных, которые после апоплексии потеряли способность говорить и у которых были ограниченные повреждения в левом полушарии. Сходные клинические данные Вернике и других исследователей воскресили идею локализации функций в мозгу. Примерно в тот же период теория локализации получила подкрепление в экспериментах Фрича и Гитцига, показавших, что электрическое раздражение определенных участков коры
Глава 9
Головной мозг
 вызывало у собаки совершенно определенные мышечные движения.
вызывало у собаки совершенно определенные мышечные движения.
В 1905 году Пьер Мари повторно исследовал препараты, представленные Брока; и показал, что его выводы тоже были основаны на ошибочных наблюдениях — что поражения мозга были значительно более обширными, чем думал Брока. Снова поддержку получила холистическая точка зрения. Обширные исследования Карла Лэшли, который в первой половине этого столетия изучал влияние повреждений мозга на результаты научения крыс в лабиринте, также подкрепляли холистическую точку зрения. Данные Лэшли указывали на то, что ухудшение функций зависит от общего объема разрушенной мозговой ткани, а не от повреждения специфических участков. Лэшли абсолютизировал представление об эквипотенциальности — любая часть коры, утверждал он, может принять на себя функцию любой другой. Однако даже он постепенно пришел к признанию того, что в коре есть некоторое «разделение труда».
, Сегодня вопрос состоит не в том, существует ли локализация функций в мозгу человека, а в том, насколько сильно она выражена. Один из наиболее авторитетных современных исследователей в этой области — советский нейрофизиолог А. Р. Лурия (1973) —выдвинул представление о том, что мозг организован как система функциональных единиц, координирующих сложные психические процессы. Во внутренней организации каждой единицы может существовать значительная пластичность, однако сами единицы связаны с определенными участками коры. Другие исследователи продолжают придерживаться мнения о большей или же о меньшей степени локализации.
Все, что мы знаем о поведении человека, говорит нам, что мозг — это не аморфное скопление клеток. Он должен обладать упорядоченной организацией, чтобы была возможна вся сложная деятельность человека. В этом отношении мы только начинаем понимать язык мозга. По-видимому, внутримозговая специализация связана не просто с каким-то анатомическим распределением клеток, а скорее со специфическими паттернами их разрядов'(John, 1972; Bartlett, John, 1973). Через сто лет наши нынешние представления о локализации функций наверняка будут казаться такими же грубыми, какими представляются нам сегодня френологические карты Галл я.
Именно в этой области уникальный вклад могут внести исследования ЭЭГ. Каким бы примитивным ни был в настоящее время метод регистрации электрической активности, она отражает функционирование мозга здорового человека. Фактически все наши современные представления о локализации
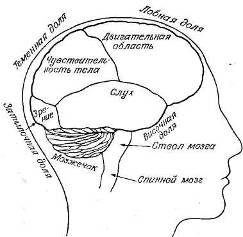
Рис. 9.2. Главные отделы мозга.
основаны на изучении животных и больных с поражениями мозга. Вводные сведения по этому вопросу превосходно изложены у Гарднера (Gardner, 1974) и Роуза (Rose, 1976). Изучение локальных ЭЭГ может расширить и дополнить имеющиеся данные.
Анатомически кору обычно подразделяют на четыре главных отдела, или доли, как это показано на рис. 9.2. Во всяком случае, ясно, что разные сенсорные модальности представлены в этих долях по-разному.
Затылочная кора участвует в обработке зрительных стимулов. Именно от нее психофизиологи чаще' всего отводили ЭЭГ, так как альфа-ритм обычно имеет здесь наибольшую амплитуду. Височная кора больше связана со слуховой системой. Поэтому, если нас интересует реакция ЭЭГ на серию тонов, то лучше будет, видимо, использовать височное, а не затылочное отведение. Соматосенсорная кора занимает небольшую область вокруг центральной борозды — углубления, разделяющего лобную и теменную доли. Именно сюда поступает осязательная информация от всего тела и отсюда посылаются двигательные команды к мышцам.
Левая и правая стороны мозга несколько различаются. Хотя многое дублируется в обеих половинах мозга, в целом наша нервная система основана на принципе перекрестной иннервации: левое полушарие мозга управляет правой половиной тела, а правое — левой. Поэтому можно ожидать, что при сжимании правого кулака изменения ЭЭГ будут более заметными в левом полушарии, и наоборот. Так оно и есть на самом
Глава 9
Головной мозг
 деле. Наконец, между функциями левого и правого полушарий существуют еще различия в психологических функциях. У правшей, например, лингвистические функции, как и предполагал Брока, связаны с левым полушарием. Мы вернемся к этому позже, когда будем рассматривать новейшие данные о межполушарной асимметрии.
деле. Наконец, между функциями левого и правого полушарий существуют еще различия в психологических функциях. У правшей, например, лингвистические функции, как и предполагал Брока, связаны с левым полушарием. Мы вернемся к этому позже, когда будем рассматривать новейшие данные о межполушарной асимметрии.
Существует много данных в пользу того, что изменения ЭЭГ, регистрируемые с различных участков головы, отражают активность соответствующих областей мозга. Например, Гасто (Gastaut, 1952) изучал в сенсомоторной области коры то, что теперь называют сенсомоторным ритмом. Этот ритм по форме похож на альфа-ритм затылочной области. Однако сенсомоторный ритм блокируется, когда человек шевелит конечностями, в то время как затылочный альфа-ритм в большей степени реагирует на зрительное внимание. Таким образом, хотя мы еще не знаем в точности механизм генерирования ритмов ЭЭГ на клеточном уровне, мы знаем, что ритмы, отводимые от разных участков черепа, обладают различной реактивностью. Есть надежда, что исследование локальных ЭЭГ поможет нам в понимании организации мозга. А изучение природы такого «биологического разделения труда» в свою очередь поможет психологам разработать биологически обоснованную классификацию форм поведения.
Расположение электродов
Мы знаем, что активность, одновременно регистрируемая с разных точек головы, может сильно различаться. Если мы поставим этот простой факт в связь с проблемой локализации, то увидим, насколько важно для электроэнцефалографии понимание основных принципов расположения электродов.
Как при всякой регистрации электрической активности, для образования замкнутой цепи следует помещать на теле два электрода. При записи ЭЭГ используются два основных метода. При биполярной записи оба электрода располагают на скальпе. В таком случае каждый из них будет улавливать электрическую активность мозга, а полученная ЭЭГ будет отражать разность потенциалов между двумя электрическими сигналами. При монополярной записи один электрод помещают на скальпе, а другой — на «электрически нейтральной» точке (чаще всего на одной из ушных мочек). На первый взгляд предпочтительнее кажется монополярная регистрация, так как можно было бы думать, что при этом отводится активность трлько одной точки мозга. Однако есть основания полагать, что биполярная запись дает нам информацию о более локальных явлениях (Lindsley, Wicke, 1974). В настоящее время относи-
тельные достоинства моно- и биполярной записи все еще остаются предметом дискуссии.
Тем не менее существует общее соглашение относительно системы точного расположения электродов. В ранних работах положение электродов часто указывали, просто называя соответствующую долю мозга, например «правая затылочная» Чтобы избежать расхождений между разными лабораториями, Международная федерация обществ электроэнцефалографии и клинической нейрофизиологии приняла систему «10—20», позволяющую точно указывать расположение электродов. Эта система представлена на рис. 9.3.
В соответствии с этой системой у каждого испытуемого точно измеряют расстояние между назионом (вдавление на переносице) и инионом (твердый костный бугорок на затылке), а также между правой и левой ушными ямками. Возможные точки расположения электродов разделены интервалами, составляющими 10 или 20% этих расстояний на черепе. Дополнительные пояснения даны в приложении Е. Для того чтобы нам можно было указывать положение электродов в разных исследованиях, отметим здесь еще несколько моментов. Буквы перед цифрами означают соответствующую долю мозга: О — затылочная доля, F — лобная, Р — теменная, Т — височная, С — область центральной борозды. Нечетные номера мест отведения относятся к левому полушарию, четные — к правому. Буквой Z обозначается отведение от верхушки черепа. Это место, которое часто используют, называется также вертексом.
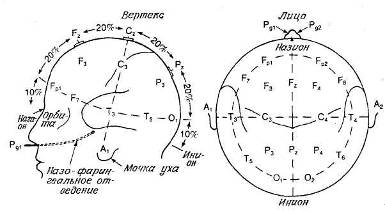
Рис. 9.3. Система 10—20. (Jasper, 1958.)
Расположение электродов на поверхности головы. F — лобная область. С — центральная, Р — теменная, Т — височная, О — затылочная. Нечетные индексы — левая половина головы, четные индексы — правая, Z — средняя линия.
144 Глава 9
Головной мозг



Рис. 9.4. Типичные электроэнцефалограммы. (Vander G., Kempe L., A Primer of Electroencephalography, Roche Laboratories, Division of Hoffman — La Roche, Inc., 1970.)
На схематических изображениях головы слева от каждой записи показано расположение электродов. Две первые записи сделаны одновременно у здорового испытуемого в состоянии покоя. Обратите внимание на высокоамплитудные альфа-волны, регистрируемые только в затылочном отведении (вверху). Третья запись —от затылочной области у здорового ребенка. У маленьких детей альфа-ритм отсутствует. На следующей записи видны необычные пики, связанные с малым эпилептическим приступом. Нижняя запись демонстрирует отсутствие электрической активности при «мозговой смерти» (хотя сердце этого больного продолжало биться, клинически он был мертв).
Применение такой системы напоминает нам, что мы не регистрируем, как это думал Бергер, «ЭКГ головы». Разные отделы мозга реагируют по-разному.
Анализ ЭЭГ
Регистрируя ЭЭГ во время сна, можно за одну ночь исписать мозговыми зигзагами больше полумили ленты полиграфа. Как свести эту гору данных к приемлемому объему?
Мы уже вкратце говорили об основных ритмах ЭЭГ (дельта, тета, альфа и бета) и о принципах частотного и амплитудного анализа. Вернемся несколько назад, чтобы рассмотреть клиническую интерпретацию ЭЭГ.
На рис.,9.4 показаны примеры нормальной и патологической ЭЭГ. В клинике, вероятно, наибольшее значение имеет
применение электроэнцефалографии для диагностики малых эпилептических припадков (petit mal). Это состояние трудно определить по поведению, поскольку его признаки — кратковременная потеря внимания, спутанность речи и т. п.— могут быть относительно слабо выражены. В таких случаях ЭЭГ особенно полезна для оценки эффективности приема больным тех или иных противосудорожных препаратов.
При записи ЭЭГ в клинике применяют достаточно стандартное расположение электродов и стандартный набор заданий, в число которых входят гипервентиляция и навязывание светового ритма, когда с помощью ритмических вспышек света искусственно вызывают судорожную активность. Поскольку при эпилепсии наблюдаются резкие изменения ЭЭГ, у клинических электроэнцефалографистов не возникало необходимости в точных критериях нормы и патологии.
Для некоторых исследователей чтение ЭЭГ стало скорее искусством, чем наукой. Воодушевленные большими успехами в выявлении аномально функционирующих участков мозга у эпилептиков, эти исследователи перешли к попыткам найти физиологические отклонения у людей с психическими аномалиями. При этом отсутствие точности в физиологическом подходе совмещалось у них с такой же неточностью при описании изменений поведения, когда были начаты исследования электроэнцефалографических коррелятов различных психозов, алкоголизма, преступности и т. п. Однако алкоголизм, психозы, преступность определялись в разных работах по-разному, а «физиологические корреляты» часто бывали просто субъективными оценками «аномальности». Не удивительно поэтому, что, несмотря на сотни таких экспериментов, не удалось прийти к какому-либо общему мнению относительно связи между психическими отклонениями и «необработанной» ЭЭГ.
В связи с этим многие исследователи ощущали необходимость в более точном подходе, чем простое рассматривание ЭЭГ. Было предложено много методов и еще больше постоянно разрабатывается. Краткое описание наиболее популярных методов дает Шагасс (Shagass, 1972), а мы ограничимся здесь лишь общим обзором возможных подходов. Сначала мы должны провести различие между методами, анализирующими одиночные и множественные отведения.
В случае одиночных отведений нас интересует изолированное рассмотрение записей, сделанных от разных отделов мозга (отведения могут быть моно- и биполярными). Для разграничения двух подходов существенно то, что в данном случае мы рассматриваем каждый раз только один канал информации. В более сложном случае — при множественных отведениях —
 |
Глава 9
мы имеем дело с фазовыми отношениями между двумя или несколькими отделами мозга, поэтому здесь необходима одновременная регистрация ЭЭГ на нескольких каналах.
И опять-таки наиболее простая форма анализа отдельных отведений — это обычный клинический метод визуального рассмотрения записей в поисках хорошо заметных особенностей. Можно достигнуть большей точности, если анализировать активность в разных частотных диапазонах. Мы можем, например, осуществлять фильтрацию ЭЭГ таким образом, что на втором канале полиграфа будет регистрироваться только активность с частотой 8—13 Гц (альфа-ри™). Как уже говорилось, мы можем считать■""какую-то минимальную ее амплитуду за «наличие альфа-ритма» и с помощью электроники или вручную определять процент времени, когда альфа-ритм (или другая полоса частот) превышает эту величину.
Более точные методы позволяют учитывать амплитуду активности в широком спектре частотных полос. Такие методы чаще всего основаны на математическом принципе рядов Фурье. Примерно 200 лет назад Фурье показал, что волна любой сложной формы математически идентична сумме синусоидальных волн разной амплитуды и частоты.
Таким образом, сложнейшие по форме волны ЭЭГ можно с помощью преобразования Фурье свести к ряду синусоидальных волн с разными амплитудами и частотами. Анализ спектра мощности дает картину относительной мощности разных частот (которая зависит от амплитуд синусоидальных составляющих). Математическое описание этих процессов очень сложно и требует применения быстродействующих цифровых вычислительных машин. Для нас сейчас важно то, что такой анализ дает значительно более полное описание ЭЭГ, нежели простой подсчет времени, которое занимает активность в том или ином частотном диапазоне. По существу, мы получаем итоговую картину распределения активности на широком диапазоне частот.
Этот метод хорошо иллюстрируют генетические исследования Ликкена и сотр. (Lykken et al., 1974) относительно ЭЭГ покоя. Производилось сравнение спектров мощности ЭЭГ у пар однояйцовых (идентичных) и двуяйцовых (неидентичных) близнецов. (Как и во многих других исследованиях по генетике человека, исходным предположением было то, что если генетически идентичные однояйцовые близнецы обнаруживают большее сходство по данному признаку, чем двуяйцо-вые, то этот признак считается наследственным.) Основной целью работы было изучение восприимчивости близнецов к гипнозу; при этом в начале и в конце одночасового эксперимента испытуемых просили сидеть три минуты с закрытыми
| Головной Most |

|
Рис. 9.5. Спектр мощности ЭЭГ у моно- и дизиготных близнецов в покое. (Lykken et al., 1974.)
Обратите внимание на то, что у однояйцовых близнецов записи очень сходны, а у двуяйцовых различаются. Это говорит о том, что параметры ЭЭГ покоя находятся под сильным влиянием наследственности. Ф — средняя частота альфа-полосы; уровень альфа-ритма (величина пика <?) выражен в процентах к мощности на частоте 3 Гц.
глазами. Как видно из рис. 9.5, между однояйцовыми близнецами обнаруживается разительное сходство. В самом деле, кривые близнецов сходны друг с другом почти так же, как сходны между собой записи каждого из них в начале и в конце
I
Глава 9
Головной мозг
 опыта. Между тем в ЭЭГ двуяйцовых близнецов сходства не обнаруживается. Коэффициент наследуемости шести различных параметров ЭЭГ оказался равным в среднем 0,82 (определенный в той же работе коэффициент наследуемости сердечного ритма в покое составил 0,67).
опыта. Между тем в ЭЭГ двуяйцовых близнецов сходства не обнаруживается. Коэффициент наследуемости шести различных параметров ЭЭГ оказался равным в среднем 0,82 (определенный в той же работе коэффициент наследуемости сердечного ритма в покое составил 0,67).
Однако вернемся к обзору методов анализа. Использование вычислительных машин позволяет осуществить еще ряд подходов к анализу относительного уровня активности в разных диапазонах частот. Из них чаще всего применяются периодический анализ (или «метод пересечения нуля») и метод автокорреляции. Однако наиболее популярными методами изучения ЭЭГ остаются применение фильтров для разных частот и анализ спектра мощностей.
При анализе множественных отведений применяются гораздо более сложные приемы. Это порождает некоторые надежды, так как мы знаем, что «трескотня» нескольких миллиардов нервных клеток, координирующих наши психические процессы,— это сложное явление, требующее комплексного подхода. Если бы ЭЭГ состояла из ряда прямых линий, ее, возможно, было бы намного легче интерпретировать, но тогда она не обещала бы нам помощи в нашем стремлении приоткрыть окно в нашу внутреннюю жизнь.
Одним из самых перспективных методов анализа множественных отведений является метод, разработанный Ливановым и его сотрудниками (1966). Записывая ЭЭГ одновременно по 50 каналам, они вычислили 1225 коэффициентов корреляции (между всеми возможными парами электродов), чтобы установить, какие области мозга работают согласованно, т. е. в каких именно участках ЭЭГ относительно синхронна. Приведем один из важнейших результатов: у здорового человека в состоянии покоя ЭЭГ большинства областей коры не синхронна, тогда'как во время решения в уме арифметической задачи активность многих корковых областей, особенно лобной коры, синхронизируется.
Технические сложности при проведении одновременной записи по 50 каналам и анализе всех возможных корреляций должны быть поистине колоссальными. Однако весьма вероятно, что для успеха в понимании скрытых функций человеческого мозга потребуется именно такого рода смелость, решимость охватить всю эту сложность, вместо того чтобы отступить перед ней.
Сон
У здорового человека только циклические изменения сна легко выдали электроэнцефалографисту тайну своего суще-
ствования. Около трети жизни (т. е. 23 полных года, если мы проживем до 69 лет) мы проводим во сне. Однако эта столь ' значительная часть нашей жизни привлекла серьезное внимание ученых лишь сравнительно недавно — в результате прогресса электроэнцефалографии (и регистрации движений глаз, как уже говорилось в гл. 7).
Описывая сон с быстрыми движениями глаз (БДГ), мы упоминали о том, что существует несколько типов сна. Там, где случайный наблюдатель в течение восьми часов будет видеть только неподвижное тело, электрофизиолог сможет построить график переходов из одной фазы сна в другую. Еще в 1862 году Кольшюттер показал, что громкость звука, способного разбудить спящего человека, систематически изменяется в течение ночи. Но для того чтобы открыть четыре стадии сна, нужно было дождаться эры электроэнцефалографии.
На рис. 9.6 показана типичная запись затылочной ЭЭГ в четырех стадиях сна, описанных Дементом и Клейтманом (Dement, Kleitman, 1957).
Засыпая, человек обычно последовательно проходит через эти стадии. Любая помеха, например неожиданный шум, может повернуть процесс вспять, возвратить спящего назад на более раннюю стадию. В течение ночного сна человек совершает переходы между этими стадиями туда и обратно,
Стадия 1
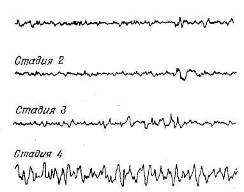
Рис. 9.6. Электрофалографические стадии сна.
В стадии 1, наступающей сразу, как только человек переходит ко сну, альфа-активность постепенно сменяется низковольтными изменениями потенциала. «Сонные веретена» — нерегулярные вспышки активности частотой 12—14 Гц — впервые появляются в стадии 2. Эти веретена вместе с высоковольтными дельта-волнами, достигающими 300 мкВ, наблюдаются и в стадии 3. И наконец, в стадии 4 в ЭЭГ преобладают высоковольтные дельта-волны.
Глава 9
Головной мозг
 периодически погружаясь в стадию БДГ со сновидениями, которая по типу ЭЭГ очень сходна с 1-й стадией. У каждого человека есть свои особенности в смене этих стадий.
периодически погружаясь в стадию БДГ со сновидениями, которая по типу ЭЭГ очень сходна с 1-й стадией. У каждого человека есть свои особенности в смене этих стадий.
Есть некоторые указания на то, что эти стадии сна функционально различны. Например, Люс и Сегал (Luce, Segal, 1966) высказывают предположение, что 4-я стадия, наиболее характерная для первых часов сна, выполняет функцию восстановления физических сил. Если вы целый день грузили ящики, то вы скорее всего проведете в 4-й стадии больше времени, чем если вы весь день читали «Введение в психофизиологию». Внешние различия между физиологически столь разными периодами сна продолжают привлекать внимание исследователей (Snyder, Scott, 1972).
Изменения ЭЭГ были обнаружены и при лишении сна. Если вы всю ночь просидели за подготовкой к экзамену, у вас, вероятно, будут короткие периоды отключения внимания («микросон»). В те несколько секунд, на которые вы отключаетесь, ЭЭГ принимает вид, характерный для спящего человека (Liberson, 1945). В этом смысле ЭЭГ оказывается индексом «умственной активации» человека.
Именно такая отчетливая смена фаз в ЭЭГ во время сна пробудила в начале 50-х годов новый интерес к «теориям активации» применительно к поведению человека (см., например, Lindsley, 1951). Напомним, однако, что на протяжении всей книги при рассмотрении физиологии человека мы постоянно подчеркивали, как важно понимать особенности каждой из систем организма. Именно поэтому концепция активации представляется нам слишком упрощенной.
Date: 2016-07-22; view: 535; Нарушение авторских прав