Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Функции ядер стриопаллидарной системы
|
|
Значение стриарной системы в целом отчетливо выявляется уже в опытах со стриарными и талами-ческими животными. Стриарные животные — животные с сохраненными стриарными телами. Стриарные кошки самостоятельно могли умываться, через неделю находили и поедали пищу; таламические были неопрятны, питались искусственно; двигательная активность у стриарных была снижена, у таламических повышена.
Таламические животные в состоянии выполнять многие простые рефлексы при изолированном их вызове, но они не могут их осуществлять в нужной последовательности. Так, таламическое животное при поднесении ему пищи ко рту облизывает ее, но не захватывает в рот, жует, но не глотает, при помещении пищи на корень языка — глотает. В то лее время стриарные кошки осуществляют указанную цепь рефлексов самостоятельно. Видимо, одна из функций полосатых тел — в обеспечении необходимой последовательности реакций при реализации сложных безусловных рефлексов.
Стриарные кошки и собаки, хотя и в несовершенной форме, но все же сохраняют способность реагировать на звуковые, световые, тактильные, проприо-цептивные, вкусовые и обонятельные раздражения. Эти животные в определенной степени сохраняют способность к выполнению сложных безусловных рефлексов — пищевых, оборонительных, ориентировочных и др.
Ряд авторов показал, что на основе этих безусловных рефлексов стриарные животные способны вырабатывать новые условные рефлексы. Таким образом, одним из уровней замыкания временной связи являются базальные ганглии. Следует отметить только, что
рефлексы в этом случае отличаются диффузностью, неточностью, плохой дифференцируемостью.
Стриарные приматы выглядят, если можно так сказать, хуже, они малоподвижны, способны только сидеть, движения у них глобальны, мало дифференцированы, плохо координированы, грубы. Живут приматы без коры всего до двух недель.
Хвостатое ядро
Эффекты повреждения хвостатых ядер. Практически повредить все хвостатые ядра не удается. Наиболее эффективен химический способ выключения подкорковых структур. Этот способ основан на том, что каждая подкорковая структура имеет некоторые специфические особенности белкового обмена, нарушение его ведет к временному выключению функции.
Выключение хвостатого ядра проводилось на крысах, кошках, собаках, обезьянах. Показано, что у крыс, кошек, собак каудотомия вызывает повышение агрессивности. Собаки с удаленными хвостатыми ядрами не могли найти дорогу в экспериментальную камеру, не узнавали экспериментатора. То же самое нарушение ориентировки в окружающей обстановке отмечалось и у кошек.
Удаление хвостатых ядер у щенят в одних случаях повышало агрессивность, в других преобладали пассивно-оборонительные реакции. Но во всех случаях отмечались остановка роста, слабость процессов концентрации возбуждения и торможения. Обезьяны с поврежденными хвостатыми ядрами не могли приспосабливаться к новым ситуациям, часто впадали в ступорообразное состояние. Повреждение хвостатых ядер вызывало снижение общей возбудимости, подвижности нервных процессов.
В человеческой патологии при страдании неостри-атума отмечаются мнестические расстройства. Амнезии возникали всегда в тех случаях, когда у больных с сотрясением головного мозга наблюдались симптомы экстрапирамидной патологии. Амнезии в этом случае могли быть ретроградные, антероградные и антерохтоградные. Наиболее четко амнезии проявлялись в случаях заинтересованности неостриатума.
Особо следует остановиться на изменениях условных рефлексов при повреждении хвостатых ядер. Повреждение хвостатых ядер у крыс приводило к полному исчезновению инструментальных условных рефлексов, причем в основном нарушалась правильность чередования действий при выполнении рефлекса. У кроликов, собак условные рефлексы после двустороннего повреждения хвостатого ядра: 1) исчезали на длительный срок; 2) выработка нового рефлекса затруднялась; 3) дифференцировка если и возникала, то была непрочной; 4) отсроченные реакции выработать не удавалось. Таким образом, повреждение хвостатых ядер приводит к нарушениям условнорефлекторной деятельности, и в первую очередь тормозных условных рефлексов (УР).
Изменения отсроченных реакций при удалении лобных долей отмечены у собак. Такие же нарушения наблюдались при повреждении переднемедиаль-ной группы ядер зрительного бугра. Морфологически показано, что между хвостатыми ядрами, зрительным бугром и лобной корой имеются тесные связи. Это все позволило выделить так называемую стриоталамо-фронтальную систему, функция которой — формирование и сохранение отсроченных реакций. При повреждении хвостатого ядра общее поведение отличалось застойностью, инертностью, трудностью переключения. Например, обезьяна долго не подходила к ры-
чагу на условный сигнал, затем, подойдя, длительно нажимала его, затем, через большой промежуток времени, шла к кормушке. У обезьян степень нарушения поведения зависела от степени и места повреждения хвостатого ядра. Наиболее существенные изменения поведения были при повреждении головки. У обезьян после одностороннего повреждения хвостатого ядра условные рефлексы восстанавливались через 30-50 дней. ЛП рефлексов удлинялись, появлялись межсигнальные. Двустороннее повреждение приводило к полному торможению условных рефлексов, видимо, двустороннее повреждение приводит к истощению симметричных компенсаторных механизмов. Высказывалось мнение (Н. Н. Олешко), что восстановление условных рефлексов происходит за счет коры головного мозга. Но, видимо, этого недостаточно, ибо в этом случае и при двустороннем разрушении хвостатого ядра компенсация, хотя бы частичная, была бы возможна.
Помимо нарушений поведения при воздействиях на хвостатые ядра отмечаются расстройства движения. Многими авторами при двустороннем повреждении полосатого тела на разных животных наблюдалось безудержное стремление двигаться вперед, одностороннее повреждение приводило к манежным движениям. У собак деструкция хвостатого ядра способствовала нарушению координации движений, последовательности движений конечностей, следовательно, посылка сигналов из коры к мотонейронам не координировалась подкоркой.
У обезьян и у многих других животных резко усиливалась общая двигательная активность при повреждении хвостатого ядра, животные постоянно находились в движении, часто меняли позы. У животных после повреждения хвостатого ядра в первые часы
наблюдалась таламическая поза. После этого появлялись тонические судороги (рис. 6.1), что свидетельствует о включении ближайшей подкорки и коры, способность самостоятельно ходить. Но не восстанавливались тонкие движения пальцев. Часто отмечались непроизвольные мимические реакции, интенционные судороги, круторазмашистый тремор, миоритмия, непроизвольные движения ног при попытке захватить предмет, анизория, птоз.

Рис. 6.1. Непроизвольные насильственные движения конечностей у обезьяны, после повреждения хвостатого ядра
У людей при повреждении связей хвостатого ядра с бледным шаром и люисовым телом может наблюдаться атетозгиперкинез дистальных отделов конечностей (кисти пальцев рук). Для атетоза характерны медленные тонические червеобразные вычурные движения. Атетоз может наблюдаться в мышцах лица: неестественное чрезмерное гримасничанье, прищуривание глаз, вытягивание губ, перекашивание рта в сторону. При атетозе гипотония мышц сменяется резким повышением их тонуса.
Эффекты раздражения хвостатого ядра. Что касается эффектов раздражения хвостатого ядра, то в
литературе данные по этому вопросу очень противоречивы. Мажанди в 1841 г. при прямом раздражении хвостатого ядра не обнаружил никаких движений. Другие исследователи при раздражении полосатого тела отмечали повороты головы, мимические реакции, тонические движения лап.
В. М. Бехтерев после повреждения коры и перерождения внутренней капсулы не обнаружил никаких движений в момент раздражения хвостатого ядра. Возникало предположение, что движения при раздражении неостриатума обусловлены петлением тока на внутреннюю капсулу.
Однако Лаурсен и др., а также Н. И. Лагутина при раздражении хвостатого ядра в участках, удаленных от внутренней капсулы, наблюдали движения, а раздражение внутренней капсулы на границе с хвостатыми ядрами не вызывало никаких движений. Далее, если двигательные эффекты зависят от петления на внутреннюю капсулу, то раздражение ее должно было бы вызвать те же эффекты, что и раздражение хвостатого ядра. Однако оказалось, что в ответ на раздражение внутренней капсулы у кошек возникали простейшие движения мышечных групп, а в ответ на раздра-.жение хвостатого ядра — сложные движения поведенческого характера. Возникновение поведенческих реакций при раздражении хвостатого ядра становится понятным, если исходить из положения, что в подкорке сосредоточены механизмы сложнейших биологических реакций. Причем у животных, стоящих по эволюционному развитию выше, увеличивается разнообразие биологических реакций, которые вызываются раздражением стриарных образований.
Так, у рыб стимуляция стриатума вызывает неспециализированные реакции типа оборонительных. У птиц при этом возникают разные виды ориентировоч-
ных реакций, оборонительных, пищевых. У млекопитающих — ориентировочные, поисковые, пассивно-оборонительные, познотонические, пищевые реакции, настороженность. При раздражении хвостатых ядер у обезьян отмечались в основном реакции тормозного типа. У собак вырабатываются слюноотделительные рефлексы на прямое раздражение хвостатого ядра. У кошек эти рефлексы вырабатываются при двигательном подкреплении. У обезьян не удалось получить двигательный инструментальный рефлекс на прямое раздражение.
Раздражение хвостатого ядра тормозит движение, вызванное раздражением коры (Лаурсен), и произвольные движения; так, Лаурсен наблюдал торможение прыжков кошки при раздражении хвостатого ядра. Раздражение хвостатого ядра тормозит также пищевые реакции, агрессивное поведение. Если наносить раздражение хвостатого ядра в момент сигнального перемещения животного, то оно не запоминает этого (рис. 6,2). У человека во время нейрохирургических

Рис.-6.2. Торможение разных этапов условнорефлекторной деятельности при стимуляции (вертикальные стрелки) хвостатого ядра: С — сигнал, Р — рычаг, К — кормушка. Сплошными линиями обозначены реализующиеся реакции, пунктирными — заторможенные
операций при раздражении хвостатого ядра останавливается речь, наблюдается выпадение повторных движений, нарушение восприятия окружающих событий, больные не запоминают задаваемых вопросов и действий персонала.
При стимуляции хвостатого ядра удлиняются ЛП рефлексов, нарушается переделка условных рефлексов. Наконец, выработка их на фоне стимуляции ядра затрудняется, а угашение в условиях стимуляции хвостатого ядра становится невозможным. Раздражение головки хвостатого ядра на разных этапах УР тормозит эти этапы. Стимуляция хвостатого ядра вызывала торможение ВП в коре на соматические, зрительные, слуховые раздражения.
Ряд подкорковых структур также получает тормозные влияния со стороны хвостатого ядра. Так, стимуляция хвостатых ядер вызывала веретенообразную активность в зрительном бугре, бледном шаре, субта-ламическом теле, черном веществе и др. Особо следует остановиться на тормозных взаимоотношениях хвостатого ядра и бледного шара. По гипотезе Тильни Рей-ли хвостатое ядро возникает эволюционно позже бледного шара. У амфибий хвостатое ядро выполняет функции торможения движений, которые присущи пал-лидарным животным. Например, рыбам для раздражения ритмических движений плавников. И действительно, удаление хвостатого ядра сопровождается усилением электрической активности бледного шара, гиперактивностью животного, раздражение хвостатого ядра снижает активность бледного шара.
Таким образом, многочисленные экспериментальные факты показали, что специфичным для раздражения хвостатого ядра является торможение активности, подкорки, торможение безусловно- и условно-рефлекторного поведения.
В то же время при раздражении хвостатого ядра могут появляться некоторые виды изолированных движений. Видимо, хвостатое ядро имеет наряду с тормозящими структурами и возбуждающие. Естественно предположить, что если возбуждение неостриату-ма тормозит движения, вызываемые с других пунктов мозга, то оно может тормозить и движение, вызываемое раздражением его самого. В то же время, когда стимулируются возбудительные системы, они вызывают то или иное движение. Далее, если считать, что функция хвостатого ядра заключается в обеспечении перехода одного вида движения в другое, т.е. прекращение одного движения и обеспечение нового путем установочных изолированных движений, то становится понятным существование двух функций хвостатого ядра — тормозной и возбуждающей.
Второе внешнее противоречие заключается в следующем. Известно, что раздражение хвостатого ядра тормозит УР на всех этапах, т.е. вызывает тормозное состояние. В то же время раздражение хвостатого ядра препятствовало угашению условного рефлекса, т.е. препятствовало развитию торможения. Однако данное противоречие кажущееся. Так как угашение является формой обучения, а стимуляция хвостатого ядра снижает способность животных реагировать (воспринимать раздражения), нарушает оперативную память, то становится понятным, почему в этом случае угасательное торможение не развивается, и животное не помнит того, что не было подкрепления. И, наконец, последнее противоречие. Стимуляция хвостатого ядра приводит к торможению УР. Следовало бы ожидать, что разрушение хвостатого ядра вызовет облегчение УР. Но оказалось, что разрушение хвостатого ядра также приводит к торможению УР.
Видимо, функции хвостатого ядра, не являются просто тормозными, а заключаются в создании условий перехода процессов одного действия в другое на основе механизмов оперативной памяти.
Все это касалось в общем-то эффекторной функции хвостатого ядра, влияния его на работу разных систем.
Что касается изменений в самом хвостатом ядре при разных состояниях организма, то таких данных чрезвычайно мало. Известны некоторые электрофизиологические изменения в хвостатом ядре под влиянием различных воздействий. У собак активность хвостатого ядра характеризовалась наличием медленных волн в фоне (1-6 Гц). Во время безусловного пищевого рефлекса электрическая картина была разной: в моменты пищепоиска она характеризовалась нестабильностью, чередованием возбуждения и торможения. В момент поедания отмечалась устойчивая электрическая активность, т.е. происходила синхронизация активности, что в коре отражалось появлением веретенной активности.
В покое активность хвостатого ядра характеризуется преобладанием низких частот. Изменения частоты пульса, количества выделяемой слюны не сопровождаются изменениями в электрической активности хвостатых ядер.
В хвостатом ядре конвергирует информация различных сенсорных систем. На основании этого делается вывод, что хвостатые ядра являются подкорковыми ассоциативными центрами. При предъявлении последовательно соматического и висцерального раздражений происходит торможение ВП на тестирующий стимул, если интервал между стимулами не более 800 мс. Причем если сравнить период рефрактер-ности в хвостатом ядре и сенсомоторной коре, то он гораздо больше в хвостатом ядре.
Процесс формирования условных рефлексов изменяет качественную характеристику неирональнои активности хвостатого ядра. УР изменял электрическую активность в хвостатом ядре по типу облегчения или торможения. Следовательно, сама неостриарная структура перестраивает активность соответственно условному рефлексу.
Известно, что нейроны хвостатого ядра полисенсорны. Нейрональная активность характеризуется низкой частотой, конвергенцией на нейронах раздражений различной модальности. Встречаются типы активности: тонический, пачечный, групповой.
Скорлупа
В литературе обычно отождествляется функция скорлупы и хвостатого ядра и имеются единичные работы, в которых изолированно изучалась скорлупа. В то же время следует отметить, что скорлупа эволюционно возникает раньше, чем хвостатые ядра. Уже одно это заставляет считать, что данная структура имеет свои особенности и функции.
Повреждение скорлупы. Опыты на кошках (Р. М. Ка-зарян, 1974) показали, что при повреждении скорлупы нарушались рефлексы пищепоиска, пищезахвата, однако эти нарушения быстро исчезали. Условные рефлексы после одностороннего повреждения скорлупы изменялись мало. В основном увеличивались ЛП. Если же опыт усложнялся выбором стороны подкрепления, то в этом случае нарушения были существенными. Правильность выбора стороны подкрепления снижалась на 30%. После двустороннего повреждения скорлупы искусственные условные рефлексы исчезали надолго. Новые условные рефлексы в этих случаях вырабатывались медленней, правильность выбора подкрепления была хуже, чем в случае до повреждения.
|
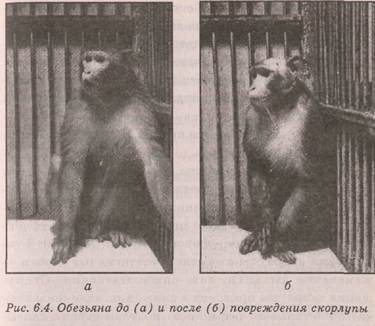
У обезьян повреждение скорлупы приводило к длительному нарушению пищевого поведения (14 дней): пищенаправленности, пищезахвата, пищеовладения (рис. 6.3). Резко снижалась агрессивность. Служители могли входить в клетку без опаски — животные стали безынициативными, вялыми. Половая активность упала. Снизилась сила нервных процессов. Животное могло часами сидеть, уставившись в одну точку (рис. 6.4). Двустороннее повреждение вызывало трофическое расстройство — язвы кожи и пр. (рис. 6.5). Нарушение пищевого поведения длилось более месяца.
Через 3 месяца после повреждения скорлупы пищевая активность резко усилилась, но овладеть пищей обезьяне было трудно из-за нарушений ВИД — пищу за решеткой на вполне доступном месте обезьяна не могла взять. Проявлялись только пассивно-оборонительные реакции, утрачивалось положение вожака. УР старые исчезли, новые не вырабатывались.

|

Рис. 6.5. Кожные трофические язвы после повреждения скорлупы
Удалось выработать только пробежку к кормущке. Дифференцировка и динамика выработки УР у обезьян с поврежденной скорлупой показали, что динамика этих процессов характерна для животных слабого
типа. ЛП длительные. Повреждение скорлупы приводило к изменению типа ВНД. Сильные животные становились слабыми. Раздражение скорлупы вызывает десинхронизацию активности в коре, подкорковых структурах. При выключении скорлупы подавлялись ВП, создаваемые раздражением кожи передних конечностей.
Эти факты объясняются активизирующим действием скорлупы на кору.
Раздражение скорлупы. Эффекты раздражения скорлупы в основном проявлялись реакциями облизывания, жевания, глотания. Эти реакции протекали, как правило, на фоне циркуляторной или ориентировочной, пассивно-оборонительной реакций. Стимуляция скорлупы вызывает вегетативные сдвиги -изменение дыхания, сна, слюноотделения. Стимуляция скорлупы приводит к снижению величины слюноотделительных рефлексов, но торможение сказывается на начале и скорости нарастания рефлекса. Стимуляция скорлупы перед условным сигналом удлиняла ЛП, стимуляция ее в момент рефлекса прекращала все его этапы, причем этот факт показан только на обезьянах. Угашение рефлекса в условиях стимуляции скорлупы затруднялось.
Следовательно, стимуляция скорлупы сказывается и на УР, и на БР (безусловных). Однако нельзя свести торможение только к БР-ному. Параллелизма величины в изменениях УР и БР не существует. При разночастотной стимуляции могло наблюдаться исчезновение УР, а БР не нарушались. В настоящее время, как уже говорилось выше, существует мнение, что хвостатое ядро и скорлупа идентичны по функции. Так, нарушения УР-деятельности (УРД) при повреждении хвостатого ядра и скорлупы однотипны — расстройство УР происходит при односторонних повреж-
дениях этих ядер на 35-40 дней, характер нарушения УРД в общем сходен, сила нервных процессов как в одном, так и в другом случае снижалась. Эффекты раздражения хвостатого ядра и скорлупы на УР были сходными. Все это предполагает единство механизмов нарушения УР при повреждении неостри-атума. Другие факты свидетельствуют о наличии функциональных различий этих структур. Так, повреждение хвостатого ядра сопровождалось повышением агрессивности и двигательной активности, а половая и пищевая активности не страдали.
При повреждении скорлупы агрессивность исчезала, преобладали пассивно-оборонительные реакции, двигательная активность снижалась, расстраивались пищевые рефлексы пищепоиска, пищенаправленнос-ти, пищезахвата, пищеовладения. Половая активность снижалась, появлялись трофические расстройства. Эти различия функций ядер имеют филогенетическое обоснование. Зачатки скорлупы возникают уже у рыб. Хвостатое ядро появляется позже, когда необходимо затормозить излишнюю двигательную активность, т.е. когда безусловные пищевые рефлексы, агрессивное поведение, половые рефлексы, вероятно, до развития хвостатых ядер в достаточной степени обеспечивались функцией скорлупы. Повреждение скорлупы приводит к возникновению у человека гиперкинезов типа торсионного спазма, хореи.
Торсионный спазм — гиперкинез шеи и туловища. Он характеризуется вращательным штопорообразным движением туловища. Хорея — быстрые изменения движения конечностей, туловища, языка, лица, мягкого нёба. Больные гримасничают, причмокивают, произвольные движения прерываются судорогами, походка напоминает своеобразный танец.
Date: 2016-11-17; view: 771; Нарушение авторских прав