Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Удивительная история эукариотических интронов
|
|
«Кусочное» (экзон-интронное) строение белок-кодирующих (и некоторых РНК-кодирующих) генов эукариот – поистине удивительная черта (она не всегда кажется нам таковой только потому, что нам так привычна концепция сплайсинга, да и открыта она, на момент написания этой книги, более 30 лет назад). Почему гены разрываются множеством некодирующих участков, большинство которых не выполняет никаких видимых функций и вырезается из транскрипта сложной молекулярной машиной (предназначенной специально для этой цели) только для того, чтобы быть разрушенными? Кажется, это превосходит все, что может нарисовать себе самое смелое воображение. Когда интроны были открыты в 1977 году, Уолтер Гилберт[66]немедленно выступил с привлекательной «гипотезой ранних интронов», послужившей основой так называемой «экзонной теории генов» (Gilbert, 1978). Вкратце, Гилберт предположил, что интроны сопутствовали жизни на самых ранних этапах эволюции и играли ключевую роль в эволюции белок-кодирующих генов, позволяя соединять короткие последовательности, кодирующие первичные пептиды, путем рекомбинации ближайших некодирующих последовательностей. Вслед за формулировкой этой идеи последовали 20 лет попыток подтвердить существование первичных интронов путем анализа разнообразных признаков интронов ныне существующих (de Souza et al., 1998). Мы не станем описывать здесь эту борьбу; достаточно лишь сказать, что убедительных свидетельств найдено не было. Безусловно, гипотеза ранних интронов не поддерживается тем фактом, что у прокариот не обнаружены сплайсосомы и интроны сплайсосомного типа, хотя Гилберт и его коллеги утверждают, что это результат эволюционного упрощения. Возможно, самый серьезный аргумент против «ранних интронов» – это обнаружение предковой связи между бактериальными самосплайсирующимися интронами и сплайсосомными интронами. Из этого открытия следует, что, даже если на самых ранних стадиях эволюции жизни существовали интроны (мы еще вернемся к этому моменту в гл. 10 и 11), эти интроны были совершенно не такими, как современные сплайсосомные интроны, и последние не могут нести какую-либо «память» о первичной эволюции. Сплайсосомные интроны и вся система сплайсинга, таким образом, суть чисто эукариотические черты из тех, что определяют «эукариотическое состояние».
Так почему же эукариотические гены прерываются таким множеством интронов? По-видимому, единственный разумный ответ следующий: потому что их предки агрессивно встраивались в эукариотические гены во время эукариогенеза или вскоре после него, а механизмы, служащие для эффективного удаления их из первичных транскриптов, развились и обеспечили выживание линии организмов со странными «кусочными» генами. После этого давление отбора, направленное на устранение интронов, во многих линиях эукариот оказалось недостаточно сильным, чтобы избавиться от большинства из них, хотя именно это произошло в других линиях, эволюционировавших в условиях сильного очищающего отбора (см. гл. 8). Разумеется, это не отменяет функциональной значимости интронов вообще: известно, что некоторые из них вносят вклад в регуляцию экспрессии (Le Hir et al., 2003), а другие даже содержат встроенные гены (Assis et al., 2008). Более того, интроны обеспечивают возможность альтернативного сплайсинга, ключевого механизма, создающего структурное и функциональное разнообразие белков у многоклеточных эукариот (см. гл. 8). В целом, однако, неискоренимое присутствие интронов, по-видимому, в большой степени зависит от силы очищающего отбора, направленного на их устранение. Популяционно-генетические аспекты утраты и приобретения интронов будут рассмотрены в главе 8; здесь я кратко обсуждаю результаты реконструкций эволюции интронов и некоторые дополнительные идеи, касающиеся природы геномов самых ранних эукариот в свете рассмотренного выше сценария эукариогенеза.
Эукариоты сильно различаются по характеру интронов: многие протисты и одноклеточные грибы содержат всего несколько интронов на весь геном, тогда как животные, растения и некоторые из простейших богаты интронами, так что кодирующие последовательности большинства их генов прерываются несколькими интронами (Jeffares et al., 2006). Замечательно, что позиции большой доли интронов консервативны у ортологичных генов разных организмов, включая растения и животных (Rogozin et al., 2003). Эволюционные реконструкции, принимающие во внимание консервативные и вариабельные позиции интронов, приводят к неожиданному выводу о том, что гены LECA были почти так же насыщены интронами, как и у современных млекопитающих, и значительная часть интронов LECA сохранилась по сей день в тех же позициях (см. рис. 7–8; Csuros et al., 2011). Этот вывод может казаться странным, но чем больше геномов становится доступно для анализа все более точными методами реконструкции, тем более убедительным он оказывается. Из этого наблюдения, формально подтвержденного результатами реконструкций, следует, что дальнейшая эволюция вела в первую очередь к утрате интронов, происходившей в большинстве ветвей эукариот, а немногие эпизоды взрывного увеличения их количества, по-видимому, были связаны с появлением новых крупных ветвей, таких как растения и животные (см. рис. 7–8). Резкое увеличение числа интронов у основания супергруппы Plantae могло быть обусловлено новой волной интронов группы II, перешедших от цианобактериального симбионта. Источник интронов у основания ветви животных остается загадочным и даже может свидетельствовать о роли скрытого эндосимбиоза в происхождении животных.
Реконструкция (см. рис. 7–8) была проведена с использованием методов Монте-Карло и марковских цепей (Csuros et al., 2011). Показана плотность интронов (число интронов на 1 Кб) для ныне существующих форм и предполагаемая плотность для ключевых предковых форм. Насыщенность черной штриховки приблизительно пропорциональна плотности интронов. Линия человека отмечена кружком. Показаны три супергруппы эукариот (Chromalveolata, Unikonta и Plantae) и основные группы внутри каждой из них, для которых известны полные последовательности геномов и соответствующие данные по локализации интронов[67].

Рис. 7–8. Реконструкция приобретения и утраты интронов в течение эволюции эукариот и плотности интронов у предковых форм.
По: Csuros et al., 2011. Данная статья находится в свободном доступе под Атрибутивной лицензией Creative Commons.
Таким образом, LECA, по-видимому, аккумулировал интроны до плотности, близкой к наиболее богатым интронами современным геномам. Что же можно сказать о динамике интронов в течение стволовой фазы, между эукариогенезом и LECA? Простой расчет показывает, что, если инвазия интронов произошла «мгновенно», то протоэукариотический геном должен был большей частью (до 80 процентов) состоять из интронов, учитывая большие и единообразные размеры интронов группы II (около 2,5 Кб; Koonin, 2009b). Скорее всего, это чрезмерное упрощение. Процесс аккумуляции интронов, вероятно, был более постепенным и сопровождался уменьшением встроившихся интронов. Как бы то ни было, интроны, по всей видимости, сыграли ключевую роль в самом начале эволюции эукариот, согласно обсуждаемой модели эукариогенеза.
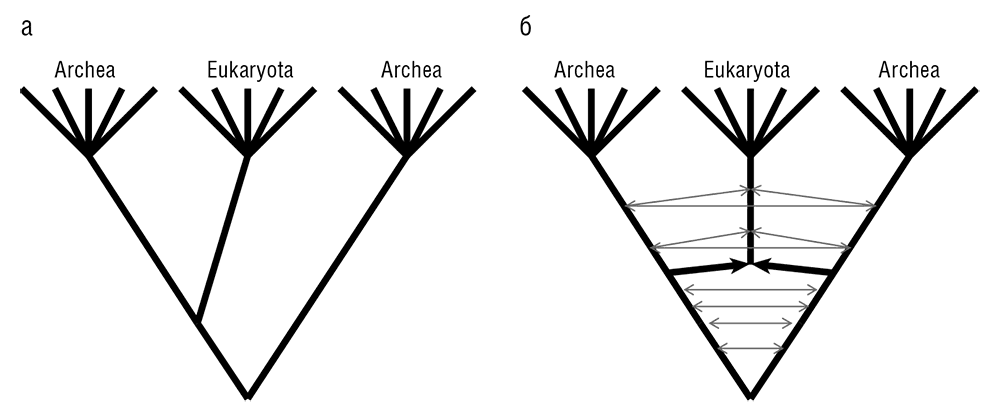
Рис. 7–9. Пересмотр трех доменов жизни: а – традиционное трехдоменное дерево Вёзе; б – циклический граф трех доменов согласно симбиогенетическому сценарию эукариогенеза и ГПГ между доменами.
Date: 2016-05-15; view: 602; Нарушение авторских прав