Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Лекция 4. Нейроны как проводники электричества
|
|
По нервам (отдельным нервным волокнам) сигналы распространяются в виде потенциалов действия и электротонических потенциалов, но на разные предельные расстояния. Способность аксонов и дендритов, а также мембран мышечных клеток проводить электрические сигналы характеризуется их кабельными свойствами.
Кабельные свойства нервных проводников очень существенны для распространения сигналов в нервной системе. Они обусловливают генерацию потенциалов действия в сенсорных нервных окончаниях, или рецепторах, при действии раздражителей, проведение сигналов по аксонам, суммацию сигналов мембраной сомы нейрона.
В основу современных теорий кабельного проведения возбуждения положена гипотеза Германна о существовании круговых токов (токов Германна), текущих от невозбужденных участков мембраны к возбужденным при распространении импульса по аксону.
Из цитологии известно, что каждая возбудимая клетка ограничена плазматической мембраной, к которой примыкают окружающие клетку оболочки. Наиболее часто нервные волокна окружены миелиновой оболочкой, сформированной глиальной клеткой в ЦНС либо оболочкой, образованной Шванновской клеткой на периферии. В местах разветвления аксона, либо в начальной или конечной его части миелиновая оболочка истончается. Сама мембрана состоит из липидов и белков. Все это обусловливает высокое электрическое сопротивление мембраны клеток и высокую распределенную электрическую ее емкость. Эти характеристики определяют проводящие свойства нервного волокна.
Основные закономерности распространения потенциалов, электротонических прежде всего, по нервным волокнам получены в экспериментах на крупных аксонах кальмаров. Было обнаружено, что при нанесении прямоугольного стимула в определенной точке волокна по мере удаления от места стимуляции сигнал регистрируется с искажением. С одной стороны, происходит изменение формы его переднего и заднего фронта (запаздывание достижения максимального значения) и уменьшение его амплитуды. Первая из этих величин определяется постоянной времени, вторая – постоянной длины. Из радиофизики известно, что постоянная времени электрической цепи имеющей емкость (С) и сопротивление R, определяется формулой
RC
и измеряется в секундах.
Из чего складывается сопротивление клеточной мембраны? В клетке существует три пути, по которым может течь ток в продольном направлении по аксону
а) аксоплазма
б)внеклеточная жидкость
в) сама мембрана
Внеклеточная жидкость –электролит, ее сопротивление мало. Удельное сопротивление мембраны толщиной 100 ангстрем приближается к 1000-5000 ом × см, очень велико. Удельное сопротивление аксоплазмы невелико, 200 ом × см. Емкость мембран возбудимых клеток С близка к 1 мкф/см2, но не бывает больше 7 мкф/см2. Таким образом, τ может быть 0,1-7 мс. Постоянная времени определяет скорость запаздывания развития потенциала до максимального значения и скорость запаздывания его затухания до фоновой величины.
Градиент нарастания потенциала (заряд мембранного конденсатора) определяется экспоненциальным законом:
V/V0=(1-e–t/τ)
Величина потенциала Vt в момент времени t меньше первоначального потенциала V0 на величину, определяемую выражением (1-e–t/τ).
Примем t=τ, тогда
Vt/V0=(1-e–1)= 1-1/e=1-1/2.7=0.63
Или 63% от первоначального.
Разряд конденсатора мембраны тоже описывается экспоненциальной формулой:
Vt/V0=e–t/τ
Примем t=τ, тогда Vt/V0=e–1=1/2,7=0,37 или 37% от максимального спустя время t.
Если через мембрану клетки течет емкостной ток электротонического происхождения, за каждый отрезок времени, равный τ, постоянной времени, сигнал электротона увеличивается на 63 % от предыдущего при возрастании сигнала, или уменьшается до 37% от предыдущей величины при его спаде.
Ионный механизм этого явления может упрощенно быть описан таким образом. При введении в клетку положительных зарядов (деполяризация) ионы К+ начинают перемещаться по направлению к мембране, которая обладает емкостью, позволяющей эти заряды накапливать, но имеются открытые каналы утечки, пропускающие ионы и демпфирующие накопление заряда. Чтобы произошел реальный сдвиг заряда, должно пройти время. Время нужно и для восстановления первоначальной величины заряда при разряде мембранного конденсатора. Это и есть τ.
На какое расстояние может распространится электротонический потенциал по мембране нервного волокна?
Пассивное распространение сигнала электротона определяется уравнением Ux=U0×e-x/λ, в котором мы снова видим экспоненциальную зависимость.
Нетрудно произвести преобразования формулы для случая x= λ и убедится, что электротонический потенциал Ux в точке, находящейся от первоначальной на расстоянии x будет меньше первоначального U0 в e раз (до 37% от U0, т.е. ½,7), если эта точка x равна постоянной длины λ.
Постоянная длины λ, или пространственная константа поляризации зависит от сопротивления мембраны rm, сопротивления внешней среды ro и сопротивления аксоплазмы ri.
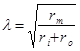
Чем больше сопротивление мембраны, меньше сопротивление среды, тем на большее расстояние передается электротонический потенциал. Уменьшение величины электротона в зависимости от расстояния называется декрементом.
Оказалось, что на постоянную длины влияет диаметр проводника, поскольку от него зависит сопротивление аксоплазмы. Поэтому толстые нервные волокна имеют большее расстояние λ, на которое способен расспространяться электротон.
Распространение электротона – прежде всего катэлектротона – важный физиологический феномен. В клетках, не генерирующих ПД (глия, эпителий, тонические мышечные волокна) за счет проведения электротона осуществляется функциональная взаимосвязь между клетками. В дендритном дереве нейронов, например, коры больших полушарий мозга, сигналы в виде электротона могут доходить от дендритов до сомы. В области синапса, зная рассмотренные закономерности, можно определить, на какое расстояние могут распространяться синаптические потенциалы.
Тем не менее, и расчеты, и прямые измерения дают очень небольшие расстояния, на которое способен распространяться электротон. Поэтому, если деполяризация в участке мембраны не достигает точки КУД, нет увеличения проницаемости для Na+, через потенциалзависимые каналы, происходят только пассивные изменения мембранного потенциала. Константа λ варьирует от 0,1 до 5 мм. Совершенно очевидно, что для связи в пределах ЦНС нужен другой механизм передачи сигналов. Эволюция его нашла. Этот механизм – распространение импульса.
Мерой проведения импульса по аксону является скорость. Скорость передачи потенциалов действия играет существенную роль в организации связей в нервной системе. Обычно быстропроводящие нервные волокна со скоростью проведения более 100 м/с обслуживают быстрые рефлексы, те, при осуществлении которых требуется безотлагательная реакция. Например, при неудачной постановке конечности (Вы оступились), чтобы избежать падения; в защитных рефлексах, инициированных повреждающим стимулом, и т.д. Для рефлексов быстрого реагирования требуется высокая скорость в афферентном и двигательном звене, до 120 м/с. Наоборот, некоторые процессы не требуют столь быстрого реагирования. Это относится к механизмам регуляции деятельности внутренних органов, где достаточно бывает скорости проведения около 1 м/с.
Рассмотрим события, связанные с распространением ПД по немиелинизированному нервному волокну (иногда с ошибкой говорят не миелиновому, но это бессмысленно, так как миелин не проводит ток, это оболочка!). Инициация волны возбуждения может быть обусловленной либо активностью рецептора (генераторный потенциал), либо синаптическими процессами. Можно вызвать ПД и электростимуляцией аксона. Если локальное смещение мембранного потенциала (деполяризация) превышает порог, достигает точки КУД, активирует потенциалзависимые натриевые каналы, ПД в виде волны деполяризации-реполяризации возникает и распространяется вдоль нервного волокна. Точка максимума ПД соответствует максимальной реверсии мембранного потенциала (овершут). Получается ситуация, при которой ПД, распространяясь по волокну, порождает перед собой токи Германна, которые разряжают мембранную емкость, приближают мембранные потенциал следующего участка мембраны к КУД, и т.д. Сзади за собой перемещающийся ПД оставляет область мембраны, находящуюся в состоянии относительной рефрактерности.
Для распространения ПД необходимо, чтобы он всякий раз порождал электротонический потенциал в соседней области, той, куда он распространяется, способный сместить мембранный потенциал на величину порога, то есть амплитуда потенциала действия должна превышать порог его возникновения в несколько раз. Отношение ПД/порог носит название гарантийный фактор (Uпд/Uпорог=5..7).
Скорость перемещения электротона и ПД по безмякотным волокнам мала и не превышает 1 м/с. У кальмара, за счет объединения в эмбриогенезе нескольких аксонов в один, что увеличивает общий диаметр проводника, скорость импульса в немиелинизированном волокне может быть до 25 м/с. У млекопитающих скорость повышена за счет миелинизации аксонов. Высокое удельное сопротивление миелина приводит к тому, что мембрана мякотных волокон приобретает высокое сопротивление и малую емкость. В перехватах Ранвье сосредоточены натриевые потенциалзависимые каналы, в приперехватных областях – калиевые, ответственные за реполяризацию. Эти особенности строение приводят к тому, что сальтаторное проведение возбуждения имеет высокую надежность и высокую скорость, которые сочетаются с экономичностью (мякотные аксоны для перемещения Na+ и K+ через мембрану обходятся меньшим количеством натрий-калиевой АТФазы). Отличительное биофизическое свойство сальтаторного проведения ПД состоит и в том, что токи замыкаются через межклеточную среду, имеющую низкое сопротивление, при этом токи следуют и вдоль, и поперек волокна.
Скорость передачи импульсов по мякотному волокну зависит от диаметра последнего простым соотношением
V=К × d, где d –диаметр, а к-константа.
Для амфибий к=2, для млекопитающих к=6.
Длина участка волокна, вовлеченного в процесс передачи одного ПД равна L=t × V, где t-длительность импульса. Этот показатель важен в методическом отношении, поскольку от длины возбужденного участка нерва зависит подбор межполюсного расстояния отводящих (регистрирующих) электродов.
В нервных стволах отдельные афферентные и двигательные нервные волокна расположены в компактно упакованном состоянии. Проведение по отдельным волокнам осуществляется изолированно от соседних, может распространяться в двух направлениях от места возникновения, имеет относительно постоянную скорость в любом участке аксона (кроме окончаний) и возбуждение от нескольких источников возникновения в клетке может подвергаться алгебраическому суммированию. Диапазон различий в скоростях проведения в волокнах велик, что позволило провести несколько классификаций. Наиболее принятыми считаются классификации Эрлангера-Гассера (группы АαβγδВС) и, в меньшей степени, Ллойда (группы I, II, III).
Date: 2015-05-23; view: 624; Нарушение авторских прав