Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Оценка метаболической функции
|
|
В клинической практике существуют приемы оценки той или иной функции:
Участие в углеводном обмене оценивается:
по концентрации глюкозы крови,
по крутизне кривой теста толерантности к глюкозе,
по "сахарной" кривой после нагрузки галактозой,
по величине гипергликемии после введения гормонов (например, адреналина).
Роль в липидном обмене рассматривается:
по уровню в крови триацилглицеролов, холестерола, ЛПОНП, ЛПНП, ЛПВП,
по коэффициенту атерогенности.
Белковый обмен оценивается:
по концентрации общего белка и его фракций в сыворотке крови,
по показателям коагулограммы,
по уровню мочевины в крови и моче,
по активности ферментов АСТ и АЛТ, ЛДГ-4,5, щелочной фосфатазы, глутаматдегидрогеназы.
Пигментный обмен оценивается:
по концентрации общего и прямого билирубина в сыворотке крови.
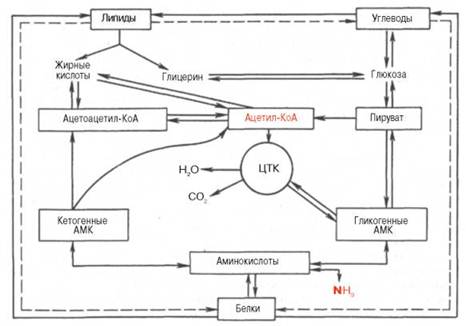

Взаимосвязь белков, жиров и углеводов.
Продукты гидролиза пищевых и тканевых триацилглицеролов, в частности высшие жирные кислоты, участвуют непосредственно в образовании сложных белков – липопротеинов плазмы крови. В составе липопротеинов, являющихся, таким образом, транспортной формой жирных кислот, они доставляются в органы-мишени, в которых жирные кислоты служат или источником энергии (сердечная и поперечно-полосатая мускулатура), илипредшественниками синтеза тканевых триацилглицеролов с последующим их отложением в клетках ряда органов (депо липидов).
Получены доказательства синтеза глюкозы из большинства аминокислот. Для некоторых аминокислот (аланин, аспарагиновая и глутами-новая кислоты) связь с глюконеогенезом является непосредственной, для других она осуществляется через побочные метаболические пути. Следует особо подчеркнуть, что три α-кетокислоты (пируват, оксалоацетат и кето-глутарат), образующиеся соответственно из аланина, аспартата и глу-тамата, не только служат исходным материалом для синтеза глюкозы, но являются своеобразными кофакторами при распаде ацетильныхостатков всех классов пищевых веществ в цикле Кребса для получения энергии.
Синтез незаменимых аминокислот из продуктов обмена углеводов и жиров в организме животных отсутствует. Клеткиживотных не содержат ферментных систем, катализирующих синтез углеродных скелетов этих аминокислот. В то же время организм может нормально развиваться исключительно при белковом питании, что также свидетельствует о возможности синтеза углеводов из белков. Процесс синтеза углеводов из аминокислот получил названиеглюконеогенеза. Он доказан прямым путем в опытах на животных с экспериментальным диабетом: более 50% введенного белка превращается в глюкозу. Как известно, при диабете организм теряет способность утилизироватьглюкозу, и энергетические потребности покрываются за счет окисления аминокислот и жирных кислот. Доказано также, что исходными субстратами для глюконеогенеза являются те аминокислоты, распад которых сопровождается образованием прямо или опосредованно пировиноградной кислоты (например, аланин, серин, треонин и цистеин). Более того, имеются доказательства существования в организме своеобразного циклического процесса – глюкозоаланинового цикла, участвующего в тонкой регуляции концентрации глюкозы в крови в тех условиях, когда в период между приемами пищи организм испытывает дефицит глюкозы. Источниками пирувата при этом являются указанные аминокислоты, образующиеся в мышцах при распаде белков и поступающие в печень, в которой они подвергаются дезами-нированию. Образовавшийся аммиак в печени обезвреживается, участвуя в синтезе мочевины, которая выделяется из организма. Дефицит мышечных белков затем восполняется за счет поступления аминокислотпищи.
Энергетическая ценность пищи оказывает определенное влияние на белковый обмен, контролируемый азотистым балансом. Так, если потребляемая энергия пищи ниже минимального уровня, то наблюдается увеличение экскрецииазота, и, наоборот, при увеличении энергетической ценности пищи экскреция азота с мочой снижается.
Между циклом лимонной кислоты и орнитиновым циклом мочевино-образования имеются сложные связи, определяющие в известной степени скорость реакций, зависимую от энергетических потребностей клетки иконцентраций конечных продуктов метаболизма. Как было показано (см. главу 12), фумаровая кислота образуется в процессе распада аргинино-янтарной кислоты, синтез которой в свою очередь требует наличия аминокислотыаспартата. Образовавшаяся фумаровая кислота (из предшественника аминокислоты аспартата) далее вступает в цикл лимонной кислоты и под действием двух ферментов этого цикла: фумаратгидратазы и малат-дегидрогеназы – превращается в оксалоацетат, который при участии специфической трансаминазы вновь превращается в аспартат, т.е. получается своеобразный аспартат-аргининоянтарный шунт цикла лимонной кислоты, соединенного с циклом мочевинообразования (рис. 15.2). Таким образом, при помощи этого необычного сцепленного механизма происходит переплетение реакций обоих циклов (мочевинообразования и ди- и трикар-боновых кислот). Этот механизм получил название «велосипед Кребса» (The "Krebs bicycle").
Из приведенной общей схемы (см. рис. 15.1) видно также, что имеются различные пути взаимопревращений жиров иуглеводов. Практика откорма сельскохозяйственных животных давно подтвердила возможность синтеза жиров изуглеводов пищи. С энергетической точки зрения, превращение углеводов в жиры следует рассматривать как накопление и депонирование энергии, хотя синтез жира сопровождается затратой энергии, которая вновь освобождается при окислении жиров в организме. Глицерин, входящий
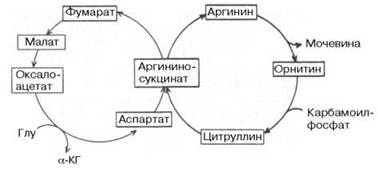
Рис. 15.2. The "Krebs Bicycle". (Печатается с любезного разрешения д-ра David L. Nelson и д-ра М.М. Сох, 1993.)
в состав триацилглицеролов и фосфоглицеринов, может легко образоваться из промежуточных метаболитовгликолиза, в частности из глицераль-дегид-3-фосфата. Следует, однако, подчеркнуть, что основным путем превращения углеводов в жиры является путь образования высших жирных кислот из ацетил-КоА, который образуется при окислительном декар-боксилировании пирувата. Последняя реакция практически необратима, поэтому образования углеводов из высших жирных кислот почти не происходит. Таким образом, синтез углеводов изжиров в принципе может происходить только из глицерина, хотя в обычных условиях реакция протекает в обратную сторону, т.е. в сторону синтеза жиров из глицерина, образующегося при окислении углеводов. Ацетил-КоА, образующийся в процессе обмена углеводов, жиров и ряда аминокислот, служит пусковым субстратом как для синтеза жирных кислот (а следовательно, и липидов вообще), так и для цикла трикарбоновых кислот. Для окисленияацетил-КоА в этом цикле требуется оксалоацетат, который является вторым ключевым субстратом в цикле Кребса. Оксалоацетат может синтезироваться из пировиноградной кислоты и СО2 благодаря реакции карбокси-лирования или образоваться из аспарагиновой кислоты в процессе транс-аминирования с α-кетоглутаратом. Две молекулыацетил-КоА, конденсируясь, образуют ацетоуксусную кислоту (ацетоацетат), которая является источником других кетоновых тел в организме, в частности β-оксимасляной кислоты (β-оксибутирата) и ацетона (см. главу 11). Следует подчеркнуть, что ацетоуксусная и β-оксимасляная кислоты часто рассматриваются как транспортные формы активной уксусной кислоты, доставляющие ее для окисления в цикле Кребса в периферических тканях. Эти жереакции конденсации двух молекул ацетил-КоА составляют начальные этапы синтеза холестерина, в свою очередь являющегося предшественником гормонов стероидной природы, витамина D3, а также желчных кислот. Последние в виде парных желчных кислот выполняют важную функцию эмульгаторов при переваривании липидов пищи в кишечнике, а также функцию транспортеров, способствуя всасыванию высших жирных кислот. Следует указать также на использование галактозы и частично глюкозы для биосинтеза цереброзидов и гликолипидов, выполняющих важные и специфические функции в деятельности ЦНС. В этом синтезе участвуют не свободные моносахариды, агексозамины (галактозамин и глюкозамин), биосинтез которых в свою очередь требует доставки амидного азотаглутамина, интегрируя тем самым обмен углеводов, липидов и белков.
В последние годы накоплено немало экспериментальных данных, свидетельствующих о существовании в живыхорганизмах множества регулирующих механизмов, осуществляющих метаболический контроль и обеспечивающих как взаимопревращения белков, липидов и углеводов, так и интеграцию энергии. Не отрицая значение других типов регуляции метаболизма (см. главы 8, 9), следует подчеркнуть, что движущей силой во взаимопревращениях веществи интенсивности метаболизма, вероятнее всего, является энергетическое состояние клетки, в частности уровень АТФ(точнее, отношение АМФ/АТФ). Так, при низких концентрациях АМФ и высоких концентрациях АТФ (состояние, которое принято обозначать «энергонасыщенностью») в клетках происходит резкое снижение глико-литического распада глюкозы, обусловленное действием этих нуклеотидов на ключевой фермент гликолиза – фосфофруктокиназу и на фосфатазу фруктозо-6-фосфата. В результате в клетках накапливается не только фруктозо-6-фосфат, но и его предшественник – глюкозо-6-фосфат. Последний, являясь положительным модулятором фермента гликогенсинтазы, стимулирует синтез полисахарида – гликогена. При низких концентрацияхАТФ (соответственно при высоком уровне АМФ) в клетках отмечаются стимулирование гликолиза и окислениепирувата в лимоннокислом цикле, что способствует обеспечению клеток энергией. Однако при низких концентрацияхАМФ имеет место снижение скорости цикла трикарбоновых кислот, обусловленное торможением активностиизоцитратдегидрогеназы, соответственно наблюдается снижение скорости синтеза АТФ и накопление изолимонной кислоты. Последняя, как известно, повышает активность другого фермента – ацетил-КоА-карбоксилазы, которая в свою очередь катализирует I стадию превращения ацетил-КоА в жирную кислоту. Благодаря этим обстоятельствамклетка переводит образовавшуюся при гликолизе молекулу ацетил-КоА с энергетического пути на путь синтезалипидов и их отложения в депо. В то же время при восстановлении скорости утилизации АТФ, что обычно наблюдается при синтезе жирных кислот, соответствующее повышение уровня АМФ способствует снижениюконцентрации лимонной кислоты и соответственно торможению синтеза липидов.
Перечисленными примерами абсолютно не исчерпывается все многообразие взаимопревращений органическихвеществ, которые постоянно совершаются в живых организмах. Здесь приведены лишь главные, магистральные каналы и пути превращения общих классов веществ и указаны ключевые субстраты и ферментные системы, обеспечивающие постоянство химических компонентов и тканей и динамичность живых структур.
Таким образом, скорость распада одних питательных веществ и биосинтеза других прежде всего определяется физиологическим состоянием и потребностями организма в энергии и метаболитах. Благодаря динамичности и координации метаболической активности обеспечивается макро- и микроскопическое постоянство всех форм живого. Выяснение фундаментальных проблем структуры и функций отдельных биомолекул может служить основой для раскрытия как молекулярных механизмов химических процессов, лежащих в основе состава и функций отдельныхклеток и целостного организма, так и процессов, обеспечивающих биологическую индивидуальность живыхорганизмов. Любые нарушения этого динамического статуса организма сопровождаются развитием патологии, тяжесть и продолжительность которой будут определяться степенью повреждения структуры и функций отдельных молекулярных и надмолекулярных компонентов клеток.
Date: 2015-05-22; view: 678; Нарушение авторских прав