Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Способы разделения и очистки белков
|
|
Получение белков из биологического материала (тканей, органов) требует такие последовательные стадии как: дробление биологического материала и разрушение клеточных мембран; фракционирование органелл, содержащих те или иные белки; перевод белков в растворённое состояние; разделение смеси белков на индивидуальные белки. Для разрушения биологического материала используют методы: создание однород. структуры в ткане, метод попеременного замораживания и оттаивания, а также обработку клеток ультразвуком. Создание однород. структуры в ткане- определ. ткань помещают в сосуд с пестиком, пестик вращается и измельчает и растирает ткань о стенки сосуда. Замораживания и оттаивания ткани - в результатеобразующиеся кристаллы льда разрушают оболочки клеток.После разрушения ткани нерастворимые части осаждают центрифугированием. Удаление из раствора небелковых веществ. Нуклеиновые кислоты, липиды и др. небелковые вещества можно удалить из раствора, используя их свойства. Липиды легко удаляются из р-ра добавлением орган-ких растворителей, например ацетона. Методы очистки белков. Очистка белков от других белков, находящихся в р-ре, полученном из данной ткани. Т.к. белки обладают лабильностью, при работе с белками следует избегать денатурирующих воздействий, поэтому выделение и очистка белков происходят при низких температурах. Также нужно учитывать термостабильность и устойчивость в кислых р-ах. Сначала нужно удалить из р-ра основную массу балластных белков, которые значительно отличаются от выделяемого белка свойствами.
13 (26). Денатурация и ренатурация. Молекулярная гибридизация нуклеиновых кислот. Денатурация свойственна макромалекулам, имеющим пространственную организацию. Она может быть вызвана нагреванием, воздействием химических веществ, которые нарушают Ван-дер-Вальсовы взаимодействия, разрывают водородные связи, стабилизирующие вторичную и третичную структуры. При нагревании происходит разделение двунитевой ДНК на одиночные полинуклеотидные цепи. При медленном охлаждении они снова воссоединяются по принципу комплементарности, образуются двунитевая полинуклеотидная цепь ДНК – этот процесс называется ренатурацией и при быстром охлаждении она не происходит. Денатурация бывает полной и частичной, обратимой и необратимой. Необратимой будет при разрушении фосфодиэфирных связей между нуклеотидами. Гибридизация нуклеиновых кислот. Все клетки одного организма содержат абсолютно одинаковые молекулы ДНК. Поэтому, если из клеток разных тканей выделить ДНК, денатурировать её и разделить цепи, а потом смешать одноцепочечные молекулы, то одноцепочечные молекулы, выделенные из разных клеток могут полностью комплиментарно спариться- образовать совершенный гибрид ДНК-ДНК.В том случае, если смешиваются ДНК разных организмов, то степень их комплим. спарив. будет зависеть от степени родстатва организмов или видов-образуются несовершенные гибриды ДНК-ДНК.
14 (12).Понятие о ферментах. структурно-функциональная организация ферментов. Ферменты-белки обладающие каталит. активностью, способны ускорять биохим-е р-ции. По своему строению могут быть простыми и сложными белками. Простетическую (небелковую) группу ферментов называют коферментом, а белковую — апоферментом. Апоферментная и коферментная группы сами по себе неактивны, лишь соединенные друг с другом более или менее прочными связями они образуют функционально активный комплекс, получивший название холофермента. Апоферментом определяется специфичность фермента (способность его взаимодействовать только с определенным веществом (субстратом) или группой веществ); тогда как коферментная группа часто выступает в роли акцептора тех или иных отщепляемых от субстрата атомов или функциональных групп, а в некоторых случаях выполняет и каталитическую функцию. Структурно-функц. организация ферментов. Центры ферментов: молекула фермента взаимодействует с S не всей своей поверхностью, а определенными участками. На поверхности фермента различают: Активный центр (это участок фермента, который взаимодействует с S. Наиболее часто в состав актив. центров входят функциональные группы таких аминокислот: ОН – группы серина, треонина, тирозина; SН – группы цистеина; NН – группа гистидина; СООН – группы глутамата и аспартата;NН2 – группы аргинина и лизина.В сложных ферментах в активный центр входят кофакторы (небелковые компоненты): простетические группы, коферменты, ионы металлов. Активный центр является комплементарным к строению S, имеется соответствие Е и S как “ключа и замка”. В структуре активного центра выделяют: участок, который связывается с S: контактный (“якорный”) участок;каталитический участок, в состав которого входят химические группы, принимающие непосредственное участие в преобразовании S. Кроме активного центра, некоторые ферменты имеют дополнительный, регуляторный, аллостерический центр, с которым взаимодействуют аллостерические регуляторы (эффекторы, модуляторы). Аллостерические эффекторы могут быть позитивными (активаторами), которые повышают каталитическую активность фермента или негативными (ингибиторами), которые ее снижают. Активный и аллостерический центры локализуются на разных субъединицах фермента. При взаимодействии аллостерического центра с эффекторами происходят конформационные изменения активного центра фермента, что приводит к увеличению или снижению его активности. Ферменты, имеющие аллостерический центр, называются регуляторными.
15 (16). Классификация и номенклатура ферментов. Все ферменты делятся на 6 классов по типу катализируемой реакции. Каждый класс делится на несколько подклассов. Классы:1.Оксидоредуктазы; 2.Трансферазы; 3.Гидралазы; 4.Лиазы; 5.Изомеразы; 6.Лигазы (синтетазы). 1.Оксидредуктазы – ферменты, катализирующие ОВР: а) аэробные дегидрогеназы (оксидазы). Отщепляют водород от субстрата и переносят его на молекулу кислорода, б) анаэробные дегидрогеназы – катализируют перенос водорода от субстрата на любой другой акцептор, кроме кислорода, в) цитохромы – это ферменты, переносящие электроны. г) пероксидазы – гемсодержащие оксидоредуктазы. Они отщепляют водород от субстрата и переносят его на Н2О2 2. Трансферазы – ферменты, переносящие группы атомов от одного субстрата к другому. При этом один субстрат донор, а другой – акцептор.- аминотрансферазы, переносят NH2;- метилтрансферазы, переносят CH3;- фосфотрансферазы, переносят PO3H2;- ацилтрнсферазы. 3. Гидролазы – ферменты, катализирующие разрыв одинарных связей с участием воды, присоединяемой по месту разрыва связи(они принимают участие в реакциях гидролиза). Все ферменты пищеварительного тракта относятся к гидролазам: эстеразы – разрывают сложно-эфирную связь; пептидазы – разрывают пептидную связь; гликозидазы – разрывают гликозидные связи. 4. Лиазы - ферменты осуществляют разрыв углеродных связей без участия воды. Выделяют: декарбоксилазы – катализируют отщепление CO2. альдолазы – катализируют расщепление связи между атомами углерода. гидратазы – разрыв двойной связи с присоединением воды по месту разрыва двойной связи. Ферменты, отщепляющие воду – реакция дегидратации. При этом образуется двойная связь. 5.Изомеразы – ферменты, катализирующие реакции изомеризации и обеспечивающие внутримолекулярную перестройку. 6.Лигазы (синтетазы) – катализируют образование более сложных вещ-в из более простых. При этом требуется энергия из вне. Обязательно участие АТФ или других. Номенклатура ферментов: 1)Тривиальная номенклатура – названия случайные, без системы и основания, например трипсин, пепсин, химотрипсин.2) Рабочая номенклатура – название фермента составляется из названия субстрата или продукта реакции, типа катализируемой реакции и окончание –аза, например лактатдегирогеназа. 3) Систематическая, научная - L-лактат-НАД-оксидредуктаза.4) Все ферменты имеют цифровой шифр, например ЛДГ - 1.1.1.27.Первая цифра говорит о типе катализируемой реакции, указывая на номер класса. Вторая уточняет действие фермента – номер подкласса. Третья указывает природу разрываемой связи в молекуле субстрата - подподкласс. Четвёртая – порядковый номер фермента.
16 (13). Общие принципы ферментативного катализа. Отличие ферментов от неорганических катализаторов. В основу современной теории о ферм.катализе положена теория Михаэлеса и Ментена. Ведущую роль в механизме ферментативного катализа играет образование фермент-субстратного комплекса. По этой теории весь процесс катализа можно разделить на 3 этапа:1)образование фермент-субстратного комплекса. Происходит диффузия субстрата к ферменту и субстрат, в соответствии с принципом комплиментарности, связывается с активным центром фермента. Реагенты связаны водородными, ионными, гидрофобными, в некоторых случаях и ковалентными связями. Эта стадия непродолжительна, зависит от концентрации субстрата и от скорости диффузии его к активному центру. Энергия активации исходных веществ при этом изменяется незначительно. На этой стадии проявляется эффект концентрирования субстрата на поверхности фермента – эффект ориентации. 2)происходит последовательное преобразование первичного фермент-субстратного комплекса в 1 или несколько активированных. Эта стадия медленная, ее длительность зависит от величины энергии активации данной реакции. В эту стадию происходит разрыв старых связей и образование новых, при этом энергия активации значительно снижается. 3)отделение продуктов от активного центра фермента и диффузия их в окружающую среду. Эта стадия непродолжительна, ее скорость определяется скоростью диффузии продуктов в окружающую среду. Отличия между ферментами и неорг. катализаторами (общие свойства ферментов): сложность строения; высокая мощность действия; специфичность; это вещ-ва с регулируемой активностью; действуют в мягких условиях организма.
17. Кинетика ферментативных реакций. Зависимость скорости ферментативных реакций от концентрации субстрата, фермента, факторов среды (рН,температуры). Число молекул S, претерпивающих изменение в течение минуты, в расчете на единицу веса E при оптимальных условиях его действия называется молекулярной активностью E. При увеличении кол-cтва E скорость ферментативной р-ции повыш-ся до некоторого предела, который характеризуется количеством S, доступным действию E. Зависимость скорости ферментативной реакции от концентрации S может быть описана уравнением:
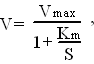
где V -скорость ферментативной реакции, Vmax - максимальная скорость ее при бесконечно большой концентрации субстрата, S - концентрация субстрата в моль/л, Кm - константа Михаэлиса (она соответствует концентрации субстрата, при которой скорость реакции равна половине максимальной). Зависимость скорости от кол-ва фермента. При условии избытка S скорость р-ции пропорциональна кол-ву E, но при избыточном количестве E прирост скорости реакции будет снижаться, поскольку уже не будет хватать S. Зависимость скорости реакции от температуры. Скорость ферментативных реакций зависит от температуры: при повышении температуры на каждые 10 °С скорость увеличивается примерно вдвое (правило Вант-Гоф-фа). Однако для ферментативных реакций это правило справедливо лишь в области низких температур — до 50-60 °С. Зависимость скорости реакции от рН. Изменение рН приводит к изменению степени ионизации ионогенных групп в активном центре, а это влияет на сродство S к активному центру и на каталитический механизм. Кроме того, изменение ионизации белка вызывает конформационные изменения молекулы E. Зависимость скорости реакции от времени. По мере увеличения времени инкубации скорость реакции снижается.Это может происходить вследствие уменьшения концентрации S, увеличения скорости обратной реакции (в результате накопления продукта прямой реакции), ингибирования E продуктом реакции, денатурации фермента. При количественном определении E и кинетических исследованиях измеряют начальную скорость реакции.
18. Уравнение Михаэлиса- Ментен. Зависимость скорости ферментативной реакции от концентрации субстрата. В 1913 г. Михаэлис и Ментен создали теорию ферментативной кинетики, исходя из того, что фермент (Е) вступает во взаимодействие с субстратом (S) с образованием промежуточного фермент субстратного комплекса (ЕS), который далее распадается на фермент и продукт реакции по уравнению:

19 (15). Механизм действия ферментов. Специфичность действия ферментов(стереохимическая, реакционная и субстратная: абсолютная, групповая). Структура и роль каталитического центра. 1. Происходит активация фермента путем связывания с аллостерическим центром регуляторных веществ это приводит к изменению конформации активного центра фермента и увеличению его способности связывать молекулу субстрата. 2. происходит 'узнавание' ферментом своего S. 3. происходит формирование неактивного фермент-субстратного комплекса за счет образования гидрофобных и водородных связей между радикалами аминокислотных остатков субстратного центра и соответствующими группировками в молекуле S. 4. образуется активный фермент-субстратный комплекс. При этом происходит хим. преобразование S с участием каталитического центра и кофермента. В результате этого молекула S меняет свою пространственную конфигурацию, в ней происходит перераспределение энергии и уменьшается прочность связей. 5. фермент-субстратный комплекс становиться нестабильным и затем преобразуется в комплекс фермент-продукт, который распадается на продукты реакции и фермент. Фермент из реакции выходит в неизменном виде.-Специфичность по отношению к субстрату – это предпочтительность фермента к субстрату определенной структуры в сравнении с другими субстратами. Различают 4 вида субстратной специфичности ферментов:1. Абсолютная специфичность – способность фермента катализировать превращение только одного S..2. Относительная специфичность – фермент катализирует превращение нескольких S, имеющих один тип связи. 3. Относительная групповая специфичность – фермент катализирует превращение нескольких S, имеющих один тип связи, но требуется наличие определенных функциональных групп, входящих в состав S. 4. Стереохимическая специфичность – фермент катализирует превращение только одного стереоизомера. Реакционная специфичность фермента -это специфичность фермента к определённой реакции. Каталитический центр - это та область актив. центра фермента, которая непосредственно участвует в хим. преобразованиях S. Формируется он за счет радикалов двух, иногда трех аминокислот, расположенных в разных местах полипептидной цепи фермента, но пространственно сближенных между собой за счет изгибов этой цепи. Если фермент явл-ся сложным белком, то в формировании каталитического центра нередко участвует простетическая группа молекулы фермента (кофермент). Коферментную функцию выполняют все водорастворимые витамины и жирорастворимый витамин K. Образование каталитического центра происходит одновременно с формированием третичной структуры белковой молекулы фермента. Чаще всего в его входят остатки серина, цистеина, тирозина, гистидина, аргинина, аспарагиновой и глутаминовой кислот.
20 (14). Кофакторы и коферменты, их значение для деятельности ферментов. Коферментные функции витаминов. Коферменты — это орган. вещ-ва аминокислотной природы, непосредственно участвующие в катализе в составе фермента. Простые, относятся обычно к классу гидролаз, практически все гидролитические ферменты состоят только изаминокислот. Кроме того, некоторые лиазы, а вот все остальные классы ферментов в основном явл. сложными белками(для каталитической активности многих ферментов кроме белковой части необходим второй компонент- кофактор). Есть каталитически актив. фермент вместе с кофактором- холофермент. Это каталитически актив. фермент, состоящий из белковой и небелковой части кофактора. Белковая часть холофермента- апофермент. У холофермента ни белковая часть апофермента, ни кофактор в отдельности не обладают заметной каталитической активностью. Оказывается апофермент резко повышает каталитическую активность кофактора, а кофактор в свою очередь стабилизирует белковую часть, делает ее более устойчивой и менее уязвимой к денатурирующим агентам. Кофакторы - это некоторые полипептиды, группы нуклеотидов и их производные и, наконец, ионы некоторых металлов. Классификация: 1) Кофакторы жирного ряда (глютатион, липоевая кислота, долихол фосфат). 2 Кофакторы алифатического ряда (убихинон или коэнзим Q). 3) Кофакторы гетероциклического ряда. 4) не содержащие витаминов - геминовые кофакторы. Основой этих кофакторов является гемовое железо. 5) Кофакторы-нуклеотиды. Коферментные функции витаминов. Ф-ции водорастворимых вытаминов-образование коферментов и простетич. групп ферментовитамины. Связанные с разл. витаминами ферменты принимают участие во мн. важнейших процессах обмена веществ: энергетич. обмене, биосинтезе и превращениях аминокислот, жирных кислот, пуриновых и пиримидиновых оснований. Некоторые жирорастворимые витамины также выполняют коферментные функции. Ф-ции др. жирорастворимых витаминов: витамин Е стабилизирует и защищает ненасыщ. липиды биол. мембран от окисления; витамин D необходим для осуществления транспорта ионов Са и остатков фосфорной кислоты через клеточные барьеры в процессах их всасывания в кишечнике, реабсорбции в почках и мобилизации из скелета.
21 (19). Регуляция активности ферментов. Ковалентная модификация. Аллостерическая регуляция. Каталитические и регуляторные центры. Понятие об иммобилизированных ферментов и их применение в медицине. Ферменты-регулируемые катализаторы.В качестве регуляторов могут выступать метаболиты, яды. Различают: активаторы – вещ-ва, увеличивающие скорость реакции; ингибиторы – вещ-ва, уменьшающие скорость реакции. Активация ферментов. Различные активаторы могут связываться либо с активным центром фермента, либо вне его. К группе активаторов, влияющих на активный центр, относятся: ионы металла, коферменты, сами S. Активация с помощью металлов протекает по различным механизмам: а)металл входит в состав каталитического участка активного центра; б)металл с S образуют комплекс; в)за счет металла образуется мостик между S и активным центром фермента.S также являются активаторами. При увеличении концентрации S скорость р-ции повышается. По достижению концентрации насыщения S эта скорость не изменяется. Если активатор связывается вне активного центра фермента, то происходит ковалентная модификация фермента: 1) частичный протеолиз. 2) фосфорилирование - дефосфорилирование. Аллостерическая регуляция. Аллостерич-е ферменты построены из двух или большего числа субъединиц. Одна субъединица имеет каталитический центр, др.— регуляторный центр. Если в среде есть аллостерический ингибитор, он присоединяется к регуляторному центру, что ведет к изменению конформации регуляторной субъединицы; вследствие этого изменяется конформация и каталитической субъединицы, в том числе каталитического активного центра. В результате активность фермента снижается. Чем выше концентрация аллостерического ингибитора, тем больше молекул фермента блокируется им и тем меньше скорость превращения субстрата. Аналогично происходит и активация ферментов при действии аллостерических активаторов. Иммобилизованные ферменты – это ферменты, ковалентно присоединенные к любому органическому или неорганическому полимеру. Их можно многократно использовать, при этом их активность снижается незначительно. При этом обеспечивается их высокая специфичность действия и высокая стабильность. В медицине иммобилизованные ферменты используются как лекарственные препараты.
22(18).Ингибирование активности ферментов: обратимое и необратимое; конкурентное, неконкурентное и бесконкурентное. Ингибитор – это вещ-во, вызывающее специфическое снижение активн-ти фермента. Инактивация –денатурация белка в результате действия денатурирующих агентов. По прочности связывания ингибитора с ферментом ингибиторы делят на обратимые и необратимые. Необратимые ингибиторы прочно связаны и разрушают функциональные группы молекулы фермента, которые необходимы для проявления его каталитической активн-ти. Обратимые игнибиторы, например прозерин для холинэстеразы. Обратимое ингибирование зависит от концентрации субстрата и ингибитора и снимается избытком субстрата. По механизму действия выделяют: 1) Конкурентное (изостерическое) ингибирование – это торможение ферментативной р-ции, вызванное связыванием ингибитора с активным центром фермента. При этом ингибитор имеет сходство с S. В процессе происходит конкуренция за активный центр: образуются фермент-субстратные и ингибитор-ферментные комплексы.2) Неконкурентное ингибирование – торможение, связанное с влиянием ингибитора на каталитическое превращение, но не на связывание фермента с S. В этом случае ингибитор может связываться и с активным центром (каталитический участок) и вне его. Присоединение ингибитора вне активного центра приводит к изменению конформации (третичной структуры) белка, вследствие чего изменяется конформация активного центра. Это затрагивает каталитический участок и мешает взаимодействию субстрата с активным центром. Неконкурентные ингибиторы: а) цианиды. Они связываются с атомом железа в цитохромоксидазе и в результате этого фермент теряет свою активность, а т.к. это фермент дыхательной цепи, то нарушается дыхание клеток и они гибнут. б) ионы тяжёлых металлов и их органические соединения. в) фармакологические средства, которые должны поражать ферменты злокачественных клеток. Сюда же относятся ингибиторы, использующиеся в сельском хозяйстве, бытовые отравляющие вещества.3) Субстратное ингибирование (бесконкурентное) – торможение ферментативной реакции, вызванное избытком S. Происходит в результате образования фермент-субстратного комплекса, неспособного подвергаться каталитическому превращению. Лекарственные препараты как конкурентные ингибиторы. Например.:четвертичные аммониевые основания ингибируют ацетилхолинэстеразу, катадизирующую реакцию гидролиза ацетилхолина на холин и уксусную кислоту. При добавлении ингибиторов активность ацетилхолинэстеразы уменьшается, концентрация ацетилхолина(S) увеличивается, что сопровождается усилением проведения нервного импульса. Ингибиторы холинэстеразы используют при лечении мышечных дистрофий.
23. Применение ферментов в медицине. Понятие об энзимопатологии, энзимодиагностике и энзимотерапии. 1. Химический метод – количественное определение S или продуктов с помощью хим. реагентов. 2. Спектрофотометрический метод – измерение скорости ферментативной реакции по изменению поглощения S при характеристической длине волны. 3. Манометрический метод – определение кол-ва газа, выделяющегося в процессе р-ции (оксидазы – по поглощению О2, декарбоксилазы – по выделению СО2). 4. Поляриметрический метод – фиксируется изменение оптического вращения. 5. Хроматографический – количественное определение S или продуктов с помощью различных видов хроматографии: бумажной (анализ сахаров), тонкослойной (гликозидов со сложными агликонами). Удельная активность – это число единиц активности (Е) на мг белка. Энзимопатология -заболевания, которые обусловлены отсутствием или снижением активности ферментов. В основном это – наследственные болезни, обусловленные генетическими нарушениями(дефекты ферментов обмена фенилаланина возникают при снижении активности фенилаланингидроксилазы-фенилпировиноградная олигофрения). Энзимодиагностика это использование определения активности ферментов в биологиче ских жидкостях для выявления тех или иных заболеваний. Энзимотерапия это использование ферментных препаратов для лечения заболеваний.
24 (22). Структурные компоненты нуклеиновых кислот. Биологическое значение и функции нуклеиновых кислот. Нуклеиновая к-та — высокомолекулярное органическое соединение, полинуклеотид, образованный остатками нуклеотидов. Нуклеиновые кислоты ДНК и РНК присутствуют в клетках всех живых организмов и выполняют функции по хранению, передаче и реализации наследственной информации. В состав нуклеиновых кислот входят азотистые основания двух типов: пуриновые - аденин (А), гуанин (G) и пиримидиновые - цитозин (С), тимин (Т) и урацил (U).Одна из основных функций нуклеиновых кислот состоит в детерминации синтеза белков. Информация о структуре белков, закодированная в нуклеотидной последовательности ДНК.
25. Вторичная и третичная структура ДНК. Типы РНК и их ф-ции. Вторичная структура ДНК -двойная спираль. Эта структура образуется из двух взаимно комплементарных антипараллельных полидезоксирибонуклеотидных цепей, закрученных относительно друг друга и общей оси в правую спираль. При этом азотистые основания обращены внутрь двойной спирали, а сахарофосфатный остов — наружу. В формировании вторичной структуры ДНК участвуют: а)водородные связи между комплементарными основаниями; б)электростатические взаимодействия; в)Ван-дер-Ваальсовы взаимодействия(связи между положительными и отриц. ионами). Третичная структура (клубок) — пространственное строение всей молекулы белка или другой макромолекулы, состоящей из единственной цепи. В формирование этой цепи участвуют: а)ковалентные связи (между двумя остатками цистеина — дисульфидные мостики); б)ионные связи между противоположно заряженными боковыми группами аминокислотных остатков; в)водородные связи; г)гидрофильно-гидрофобные взаимодействия. На поверхности молекулы оказываются полярные гидрофильные боковые группы. Типы РНК и их функции. Три типа РНК: – транспортные (тРНК), матричные (мРНК) и рибосомальные (рРНК). Они различаются по первичной структуре, молекулярной массе, конформации, продолжительности жизни и выполняемым функциям. Транспортные РНК -описывается универсальной моделью «клеверного листа». В состав тРНК входят минорные основания, представленные метилированными основаниями, изомерами и аналогами пиримидинов. Минорные основания делают тРНК устойчивыми к действию нуклеаз и поддерживают определенную третичную структуру молекулы. Антикодон имеет жесткую архитектуру, которая позволяет ему быстро считывать мРНК. Матричные РНК эукариот и прокариот различаются по строению. Этот тип РНК имеет одинаковое строение 5′- и 3′-концов.На 5′ конце присутствует модифицированный нуклеотид 7-метилгуанозин-5′-трифосфат – кэп. Рибосомальные РНК принимают участие в образовании рибонуклеопротеинов, формирующих немембранные комплексы – рибосомы. Клетки прокариот и эукариот содержат рибосомы, имеюшие общий план строения. В рибосомы входят высокомолекулярные рРНК, дающие начало 30S-40S- и 50S-60S-субчастицам рибосом; рРНК взаимодействуют с мРНК и аминоацил-тРНК в процессе трансляции. рРНК содержат несколько модифицированных нуклеотидов. Чаще всего это метильные производные азотистых оснований или рибозы. Вторичная структура рРНК характеризуется спирализацией самой на себя полирибонуклеотидной цепи. Биспиральные и линейные участки этих молекул формируют постоянные вариабельные домены, которые затем укладываются в более компактные структуры более высокого порядка.
26 (23). Строение и уровни организации нуклеиновых кислот. Первичная структура нуклеиновых кислот. Видовые различия первичной структуры нуклеиновых кислот. Нуклеиновая к-та — высокомолекулярное органическое соединение, полинуклеотид, образованный остатками нуклеотидов. В состав нуклеиновых кислот входят азотистые основания двух типов: пуриновые - аденин (А), гуанин (G) и пиримидиновые - цитозин (С), тимин (Т) и урацил (U). Уровни структурной организации: 1. первичная структура. РНК и ДНК построены однотипно – представлены полинуклеотидной цепью, состоящей из отдельных мононуклеотодов, соединённых между собой 3’→5’-фосфодиэфирными связями. 2. вторичная структура. Представляет собой двойную спираль, состоящую из 2 полинуклеотидных цепей, закрученных вокруг одной общей оси. Эти цепи антипараллельны, т.е. одна идет в направлении 5’→3’, а другая 3’→5’. Пуриновому основанию одной цепи соответствует пиримидиновое основание другой цепи – эти основания комплиментарны друг другу. Вторичная структура стабилизируется водородными связями и гидрофобными взаимодействиями. Видовые различия первичной структуры: Существуют два типа нуклеиновых кислот – дезоксирибонуклеиновые кислоты (ДНК) и рибонуклеиновые кислоты(РНК). те, и другие представляют собой полимерные молекулы, построенные из мономерных блоков – нуклеотидов: ДНК – из дезоксирибонуклеотидов, а РНК – из рибонуклеотидов.
27 (24). Вторичная и третичная структура ДНК. Строение и организация хроматина. Вторичная структура ДНК -двойная спираль. Эта структура образуется из двух взаимно комплементарных антипараллельных полидезоксирибонуклеотидных цепей, закрученных относительно друг друга и общей оси в правую спираль. При этом азотистые основания обращены внутрь двойной спирали, а сахарофосфатный остов — наружу. В формировании вторичной структуры ДНК участвуют: а)водородные связи между комплементарными основаниями; б)электростатические взаимодействия; в)Ван-дер-Ваальсовы взаимодействия(связи между положительными и отриц. ионами). Третичная структура (клубок) — пространственное строение всей молекулы белка или другой макромолекулы, состоящей из единственной цепи. В формирование этой цепи участвуют: а)ковалентные связи (между двумя остатками цистеина — дисульфидные мостики); б)ионные связи между противоположно заряженными боковыми группами аминокислотных остатков; в)водородные связи; г)гидрофильно-гидрофобные взаимодействия. На поверхности молекулы оказываются полярные гидрофильные боковые группы. Хроматин -это вещ-во хромосом — комплекс ДНК, РНК и белков. Хроматин находится внутри ядра клеток эукариот и входит в состав нуклеоида у прокариот. Именно в составе хроматина происходит реализация генетической информации, а также репликация и репарация ДНК. Основную массу хроматина составляют белки гистоны. Гистоны являются компонентом нуклеосом, — надмолекулярных структур, участвующих в упаковке хромосом. Нуклеосомы располагаются довольно регулярно, так что образующаяся структура напоминает бусы. Нуклеосома состоит из белков четырех типов: H2A, H2B, H3 и H4. В одну нуклеосому входят по два белка каждого типа. Нить ДНК с нуклеосомами образует нерегулярную структуру (фибриллу). Если хроматин упакован плотно- гетерохроматин, ДНК, находящаяся в гетерохроматине не транскрибируется. В интерфазе гетерохроматин обычно располагается по периферии ядра (пристеночный гетерохроматин). Полная конденсация хромосом происходит перед делением клетки. Если хроматин упакован неплотно- эухроматином. Плотность упаковки хроматина во многом определяется модификациями гистонов — ацетилированием и фосфорилированием.
28. Репликация ДНК как один из видов матричных синтезов. Этапы репликации. Особенности процесса в эукариотических клетках. Репликация (самоудвоение, биосинтез) ДНК. Основные этапы: 1. образование репликативных вилок при участии ДНК-раскручивающих белков, вызывающих разрыв водородных связей между комплементарными основаниями. 2. синтез новых нитей ДНК при участии ДНК-полимеразы, катализирующей образование фосфодиэфирной связи между новыми нуклеотидами. Присоединение нуклеотидов идет в соответствии с принципом комплементарности. Синтез идет 5’-конца к 3’-концу. На одной цепи синтез происходит непрерывно, а на другой - прерывается с образованием коротких фрагментов. В результате на одной цепи образуются короткие фрагменты - фрагменты Оказаки; 3. соединение коротких фрагментов с помощью ДНК-лигазы с образованием дочерних нитей. В результате репликации на одной материнской нити синтезируются 2 комплементарных дочерних ДНК (из одной молекулы ДНК образуются 2 копии ДНК). Значение репликации состоит в передаче информации от ДНК матери к дочерней ДНК. Особености репликация линейных геномов эукариот:геномы кольцевые замкнутые характерны для многих бактерий, их плазмид и некоторых вирусов. У большинства др. организмов геном представлен линейными молекулами ДНК в составе одной или нескольких хромосом. Существует так называемая проблема отстающей цепи ДНК. Синтез отстающей цепи ДНК происходит в виде коротких фрагментов Оказаки, для инициации синтеза которых требуются РНК- затравки. После удаления затравки на конце одной из вновь синтезированных молекул ДНК образуется одноцепочечная, которая не может быть заполнена ДНК-полимеразой, поскольку она не функционирует в отсутствие праймера. Вследствие этого в каждом раунде репликации должно было бы происходить укорачивание хромосом с обоих концов, что приводило бы к потере генетической информации, закодированной в концевых фрагментах ДНК. Кроме того, большие размеры молекул ДНК, заключенных в индивидуальные хромосомы, требуют специальной организации их реплицирующего аппарата.
29. Репликация плазмид. Особенности репликации вирусного генома. Интерфероны, их биологическое действие и применение в медицине. Плазмида - внехромосомный самовоспроизводящийся генетич. элемент бактерий. Представляет собой кольцевую двухцепочечную молекулу ДНК, закрученную в суперспираль. Каждая плазмида является самостоятельным репликоном, сама контролирует собственную репликацию и поддерживается в клетке в определенном, характерном для нее числе копий. Особенности репликации вир-го генома: I) вирусная частица (вирион) содержит только один вид нуклеиновых кислот — или ДНК, или РНК; 2) вирионы не имеют собственного метаболизма, не содержат клеточных органелл, в том числе рибосом, и очень часто состоят только из нуклеиновой кислоты, заключенной в белковую оболочку=>вирусы способны размножаться исключительно за счет использования метаболического аппарата другой клетки.Цикл размножения вируса начинается с его прикрепления к поверхности клетки. Вирион содержит рецепторы, узнающие определенные вещества мембраны клетки-хозяина; После прикрепления вирион проникает чз мембрану внутрь клетки; иногда в клетку попадает только нуклеиновая кислота вириона. Затем с использованием аппарата клетки-хозяина начинается репликация вирусного генома и синтез вирусных белков; из них путем самосборки образуются новые вирионы, которые освобождаются из клетки, либо разрушая ее. Интерфероны — ряд белков со сходными св-ми, выделяемых клетками организма в ответ на вторжение вируса. Они способны подавлять внутриклеточное размножение ДНК- и РНК-вирусов. В медицине интерфероны применяют для профилактики и лечения респираторных вирусных инфекций.
30 (29). Биосинтез РНК (транскрипция). Строение РНК-полимеразы. Зависимость локализации считываемого участка и направления считывания от структуры промотора. Этапы транскрипции. Посттранскрипционная модификация РНК. Процессинг РНК. Транскрипция - передача информации с ДНК на РНК. Элементарная единица транскрипции-оперон (транскриптон)- участок ДНК, подвергающийся транскрипции. В опероне выделяют информативные участки - экзоны и неинформативные участки - интроны. В начале оперона выделяют промотор (P) - это начальный участок оперона, к которому присоединяется РНК-полимераза. Рядом с промотором располагается оператор (О) - регуляторная зона, место присоединения генов-регуляторов. В конце оперона располагается терминатор (Т) - участок, содержащий стоп-сигнал. Строение РНК-полимеразы. Биосинтез РНК осуществляется ДНК-зависимыми РНК-полимеразами. В ядрах эукариотов обнаружены 3 специализированные РНК-полимеразы: РНК-полимераза I, синтезирующая пре-рРНК; РНК-полимераза II, ответственная за синтез пре-мРНК; РНК-полимераза III, синтезирующая пре-тРНК. Этапы транскрипции: 1. инициация; 2. элонгация; 3. терминация;4.процессинг (созревание) РНК. Инициация - присоединение ДНК-зависимой РНК-полимеразы к промотору, что приводит к разрыву водородных связей между комплементарными нуклеотидами и расхождение нитей ДНК. Элонгация - это передвижение РНК-полимеразы вдоль нити ДНК, сопровождающееся образованием фосфодиэфирных связей между рибонуклеотидами. Присоединение рибонуклеотидов происходит в соответствии с принципом комплементарности. Этот синтез идет от 5' конца к 3'. Данная фаза протекает до тех пор, пока ДНК-полимераза не достигнет стоп-сигнала->терминация. В результате процессов транскрипции образуется транскрипт (пре-иРНК). Он почти полностью соответствует транскриптону. Посттранскрипционная модификация РНК. Процессинг РНК. Начинается следующий этап - процессинг - посттранскриптационное созревание РНК. Это удаление излишков - вырезаются неинформативные участки; сплайсинг - сшивка, соединение информативных участков. При этом иРНК укорачивается.Из пре-иРНК образуется иРНК. Далее иРНК соединяется с белком-информером, в результате чего образуется комплекс иРНК+ белок =информосома, который может выходить из ядра и транспортироваться в цитоплазму к рибосоме, где начинается – трансляция.
31. Основной постулат молекулярной биологии. Генетический код и его характерные черты. Акцепторная роль тРНК. Синтез аминоацил-тРНК как регуляторный механизм трансляции. Основной постулат молекулярной биологии. Передача наследственной информации от материнской клетки к дочерней осуществляется при помощи ДНК-репликация. Для использования генетической информации необходимы РНК, образуемые на матрице ДНК-транскрипция. Далее РНК непосредственно участвуют на всех этапах синтеза белковых молекул трансляция, обеспечивающих структуру и деятельность клетки. Генетический код. Генетическая информация, содержащаяся в ДНК и в иРНК, заключена в последовательности расположения нуклеотидов в молекулах. Последовательность расположения нуклеотидов в иРНК определяет последовательность расположения аминокислот в белках. Этот код называют генетическим. Код триплетен. В состав РНК входят 4 нуклеотида: А, Г, Ц, У(каждая из 20 аминокислот зашифрована последовательностью 3 нуклеотидов, т. е. триплетом, который получил название кодон) Код однозначен. Каждый триплет шифрует только одну аминокислоту. Код универсален. Код един для всех живущих на Земле существ. Акцепторная роль тРНК. тРНК-осуществляет доставку аминокислот к рибосоме. Связывается с аминокислотами, образуя комплекс аминоацил-тРНК, при помощи аминоацил-тРНК-синтетазы. Синтез аминоацил-участие в регуляции. Аминоацил-тРНК-синтетаза— фермент, катализирующий образование аминоацил-тРНК в реакции этерификации определенной аминокислоты с соответствующей ей молекулой тРНК. Для каждой аминокислоты существует своя аминоацил-тРНК-синтетаза.
32 (31). Этапы трансляции. Состав трансляционного аппарата клетки. Строение и механизм функционирования рибосом. Роль РНК в процессе трансляции. Участие белковых комплексов инициации, элонгации и терминации в биосинтезе полипептидной цепи. Трансляция - перевод генетического текста иРНК в последовательность аминокислот в белке. Этапы трансляции: 1. Рекогниция. Происходит связывание АК со своей тРНК. 2. Инициация. Состоит из 7 фаз: а) подготовка рибосомы к трансляции. Рибосома взаимодействует с фактором инициации, в результате разделяется на малую и большую субъединицы. б) подготовка РНК-матрицы к трансляции; в) подготовка инициаторной аминоацил-тРНК. г) образование инициирующего комплекса. д) связывание мРНК с инициирующим комплексом. е) поиск и комплементарное взаимодействие со стартовым кодоном. ж) формирование 80S-рибосомы. В образовавшейся рибосоме выделяют 2 участка: Р-участок, там будет происходить образование пептидных связей; А-участок (аминоацильный участок) – служит для присоединения аминоацильной- тРНК. 3. Элонгация состоит из 3-х фаз: а) присоединение следующей аминоацил-тРНК в соответствии со смыслом следующего кодона матрицы; б) пептизация; в) транслокация или перемещение. Рибосома перемещается на 1 кодон в направлении 3’-конца тРНК. При достижении рибосомой нонсенс-кодона элонгация прекращается и наступает терминация. 4. Терминация. Нонсенс-кодон распознается белковыми R-факторами (факторы освобождения) в А-участке рибосом. В результате действия R-факторов обеспечивается диссоциация элементов трансляционного аппарата и в цитоплазму высвобождаются рибосома, иРНК, полипептид. Трансляция включает 3 этапа: инициация, элонгация, терминация. 5. Процессинг белка -Это совокупность изменений в структуре белка, которые заканчиваются формированием структурно и функционально зрелой белковой молекулы.он может быть 2-х видов: а)котрансляционный, протекающий во время трансляции; б)посттрансляционный – химическая модификация белка происходит после трансляции. Рибосомы. Каждая рибосома состоит из малой и большой субъединиц. Содержит рРНК, а также около 80 различных белков, в том числе и ферментов. Трансляция — синтез белка рибосомой на основе информации, записанной в матричной РНК. мРНК связывается с малой субъединицей рибосомы, когда происходит узнавание 3'-концом 16S рибосомной РНК комплементарной последовательности Шайн-Далгарно, расположенной на 5'-конце мРНК, а также позиционирование стартового кодона мРНК на малой субъединице. У эукариот малая субчастица рибосомы связывается также с помощью кэпа, на конце мРНК. Ассоциация малой и большой субъединиц происходит при связывании формилметионил-тРНК и участии факторов инициации. Таким образом, распознавание антикодона происходит на малой субъединице.
33. Регуляция биосинтеза белка на уровне репликации и транскрипции. Регуляция биосинтеза белка на этапе трансляции. Посттрансляционная модификация белков. Основные уровни регуляции биосинтеза: 1. Регуляция синтеза белка на стадии репликации. Регуляция возможна путем добавления (или удаления) кодирующих (или регуляторных) участков на ДНК. Если удалить матрицу, то и синтеза белка с нее не будет, а если увеличить дозу определенного гена в ДНК, то появится возможность ускорить построение белков на многих одинаковых матрицах. В большинстве случаев наличие избыточной генетической информации отрицательно сказывается на жизнеспособности клеток, а ее недостаток, возникающий из-за недорепликации ДНК, приводит к их гибели из-за отсутствия жизненно важных генов. 2. На уровне транскрипции.
Варианты: а)групповая репрессия генов белками – гистонами; б)амплификация генов - увеличение числа копий заданного участка ДНК или гена. в)регуляция сигналами-усилителями. Сигналы-усилители - энхансеры - выступающий участок ДНК, который может быть значительно удален от промотора. 3. регуляция на уровне трансляции: тотальная репрессия или активация трансляции при изменении активности и количества белковых факторов; избирательная дискриминация иРНК; механизм повышения эффективности трансляции включает образование полисом - это комплекс нескольких рибосом с одной иРНК. 4. Регуляция биосинтеза белков путем посттрансляционной модификации: приводила к образованию общей полипептидной цепи, разрезаемой в дальнейшем на индивидуальные белки специфическими протеиназами. Посттрансляционная модификация белков. Модификации происходят в ЭПС и аппарате Гольджи. Однако множество других ковалентных модификаций протекает в первую очередь именно в цитозоле. Некоторые из них стабильны и необходимы для активности белка. Посттрансляционные модификации включают в себя фосфорилирование факторов транскрипции протеинкиназами, гликозилирование, ацилирование, циклизацию, амидирование, гидроксилирование и метилирование различных остатков аминокислот. Многие из перечисленных модификаций являются критическими для биологической активности пептидов.
34. Теория оперонной регуляции транскрипции. Ф-ии и особые зоны промотора. В 1961 году при исследовании индукции синтеза фермента—β-галактозидазы, участвующего в расщеплении лактозы в клетках E. coli, было установлено следующее: когда клетки E. Coli растут на среде, содержащей глюкозу, то в них содержится менее 10 молекул β-галактозидазы на клетку. Если же в среде глюкозу заменить на лактозу, то ч/з несколько минут наблюдается индукция синтеза белков, и концентрация ферментов утилизации лактозы увеличивается в сотни раз. Это явление объяснила теория оперона, доказавшая, что на молекуле ДНК можно обнаружить участки — опероны, которые содержат информацию о группе функционально взаимосвязанных структурных белков, и регуляторную зону (участки оператора и гена регулятора), контролирующую транскрипцию этих генов. Экспрессия структурных генов определяется способностью РНК-полимеразы связываться с промотором, расположенным на 5'-конце оперона. Присоединение фермента зависит от оператора, участка ДНК, который находится рядом с промотором и даже частично с ним перекрывается. Оператор может связываться с белком-репрессором, который синтезируется в клетке с постоянной скоростью. Строение белка-репрессора кодирует мРНК, транскрибируемая с гена-регулятора, расположенного на определенном расстоянии от оперона, работу которого он контролирует. Если белок-репрессор присоединяется к оператору, то РНК-полимераза не может связаться с промотором и транскрипция структурных генов не идет. Белки, участвующие в утилизации лактозы, практически не синтезируются. Когда в среде появляется индуктор — лактоза и присоединяется к белку-репрессору, то последний изменяет конформацию и теряет сродство к оператору. РНК-полимераза связывается с промотором, структурные гены транскрибируются, синтезируется одна полицистронная молекула мРНК, содержащая информацию о трех белках: β-галактозидазе, пермеазе и галактозидтрансацетилазе, необходимых для утилизации лактозы клетками. При регуляции оперона по механизму репрессии белок-репрессор в клетках, выращенных на среде, содержащей NH4CL в качестве единственного источника азота, не имеет сродства к оператору. Транскрибируются структурные гены оперонов, которые кодируют ферменты. Когда добавляют корепрессор — аминокислоту, за синтез которой отвечает один из этих оперонов, то она связывается с белком-репрессором. Образуется комплекс белок-репрессор — корепрессор, который приобретает сродство к оператору, связывается с ним и транскрипция структурных генов прекращается.
35. Молекулярные механизмы генетической изменчивости. Виды и причины мутаций, связь между мутагенными факторами и типом мутаций. 1)Повреждения и репарация ДНК, 2)Апоптоз- механизм запрограмированной и регулируемой гибели клеток. Он начинается с повреждений систем репараций ДНК и накопление повреждений ДНК. Эти изменения активируют ряд специфических протеаз в клетке, которые активируют эндонуклеазы. Эндонуклеазы гидролизуют ДНК сначала на крупные фрагменты, а затем происходит гидролиз по межнуклеосомным областям ДНК и образуются фрагменты. Далее клетка распадается на мембранные везикулы, содержащие фрагментированную ДНК и другие компоненты клетки, и всё это впоследствии поглощается фагоцитирующими клетками. Таким путем устраняются клетки, размножение которых может быть опасным для организма, например привели к развитию раковой опухоли.3) Мутагенез, вследствии воздействия мутагенов(нитраты, УФ излучение и т.д.) ВИДЫ МУТАЦИЙ: Точечная мутация, или единственная замена оснований, — тип мутации в ДНК или РНК, для которой характерна замена одного азотистого основания другим. Хромосомные мутации- Хромосомные мутации - это перестройки хромосом. Участки хромосом могут изменить свое положение, потеряться или удвоиться. 1.Инверсии-поворот участка на 180градусов. 2. Транслокации - перенос какого-то участка хромосомы в другое место той же хромосомы или на другую хромосому3. Делеции- выпадения букв из цепи.4. Дупликации - тип мутаций, заключающийся в удвоении какого-либо участка хромосомы. Геномные мутации 1. Анеуплоидия (уменьшение количества хромосом в 2 и более раз)2. Полиплоидия (увеличение количества хромосом в 2 и более раз). Ядерные и цитоплазматические мутации 1. Ядерные мутации — геномные, хромосомные, точечные. 2. Цитоплазмотические мутации — связанные с мутациями неядерных генов находящихся в митохондриальной ДНК и ДНК пластид — хлоропластов.
36. Классификация мутаций. Геномные мутации. Нерепарируемые мутации и их последствия. Мутации разделяют на геномные, хромосомные и генные. При геномных мутациях (полиплоидия) у организма-мутанта происходит внезапное изменение числа хромосом, кратное целому геному. Различают аллополиплоидию, в рез-те которой происходит объединение при гибридизации целых неродственных геномов, и аутополиплоидию - увеличение числа хромосом собственного генома, кратное 2n. При хромосомных мутациях происходят как изменение числа отдельных хромосом в геноме (анеуплоидия), так и крупные перестройки структуры отдельных хромосом (аберрации). В этом случае наблюдаются потеря (делеции) или удвоение части (дупликации) генетического материала одной или нескольких хромосом, изменение ориентации сегментов хромосом в отдельных хромосомах (инверсии), а также перенос части генетического материала с одной хромосомы на другую (транслокации). При генных мутациях происходят замены, делеции и вставки одного или нескольких нуклеотидов, транслокации, дупликации и инверсии различных частей гена. Генные мутации -нерепарированные наследуемые изменения первичной структуры ДНК, которые ведут либо к прекращению синтеза белка, кодируемого поврежденным геном, либо к синтезу измененного, «неправильного» белка.
Date: 2015-09-25; view: 600; Нарушение авторских прав